
Войти
| Garudimimus. Временной диапазон: Поздний мел,. ~ 96–89 Ma PreꞒ Ꞓ O S D C P T J K Pg N | |
|---|---|
 | |
| Установленный голотип в Монгольском музее естественной истории (обратите внимание на неправильные позвоночные позвонки; в голотипе указаны только четыре) | |
| Научная классификация | |
| Домен: | Eukaryota |
| Царство: | Animalia |
| Тип: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Maniraptoriformes |
| Clade: | †Ornithomimosauria |
| Род: | †Garudimimus. Barsbold, 1981 |
| Типовой вид | |
| †Garudimimus brevipes . Барсболд, 1981 | |
Garudimimus (что означает «Garuda мимик») - это род орнитомимозавров, которые жили в Азии во времена Поздний мел. Род известен по единственному экземпляру, найденному в 1981 г. советско-монгольской экспедицией в формеции Баян Ширех официально описанным в том же году Ринченом Барсболдом ; единственный вид - Garudimimus brevipes . Несколько интерпретаций анатомических показателей Garudimimus были сделаны при последующем исследовании этого экземпляра, но большинство из них подверглось критике во время его всеобъемлющего переописания в 2005 году. Обширные неописанные останки орнитомимозавров в типовой местности Garudimimus особые экземпляры этого рода.
Единственный известный экземпляр гарудимимуса - животное среднего размера, длиной 3,5 м и весом около 98 кг (216 фунтов). Это был орнитомимозавр со смесью основных и производных черт; в отличие от примитивных орнитомимозавров, и верхняя, и нижняя челюсти были беззубыми, черта, которая часто сообщается у более производных орнитомимид. У гарудимимусов были относительно короткие и коренастые задние конечности, крепкие лапы и уменьшенная подвздошная мышца. На стопе было четыре пальца, первый из которых был сильно уменьшен, в то время как у орнитомимидов был трехпалый палец с утраченным первым пальцем. У беззубого черепа очень прямые челюсти, заканчивающиеся более закругленным кончиком морды, чем у других родов. Ранее считалось, что этот примитивный орнитомимозавр обладал слезным «рогом» на вершине черепа перед глазницей. Однако повторное описание единственного образца показало, что эта структура была просто искаженной левой префронтальной костью. Другой раннейшей интерпретацией была реконструкция плюсневой кости с использованием арктометатарсалий.
При первом описании в 1981 году Гарудимим был идентифицирован как примитивный орнитомимозавр в пределах своего собственного семейства. Однако при описании новых образцов дейнохейруса в 2014 году было обнаружено последний сестринским таксоном Garudimimus, входящим в группу Deinocheiridae - орнитомимозавров. не приспособлен для бега или подвижных движений. Но это размещение также оказалось маловероятным. Тазовый пояс и задние конечности гарудимимуса показывают, что мускулатура ног не так хорошо развита, как у быстро бегущих орнитомимидов, что указывает на слабые беглые способности. Как и другие представители Орнитомимозаврии, Гарудимим был всеядным / травоядным животным с уменьшенной силой укуса, компенсировалась роговым клювом.
 определения окаменелостей динозавров мелового периода в Монголии; Окаменелости гарудимимуса были собраны в местонахождении Байши Цав области C (Форма Баян Ширех )
определения окаменелостей динозавров мелового периода в Монголии; Окаменелости гарудимимуса были собраны в местонахождении Байши Цав области C (Форма Баян Ширех )В 1981 году во время советско-монгольской экспедиции в пустыню Гоби был обнаружен относительно скелет теропод. позднемеловом формации Баян Ширех, юго-восток Монголия. Останки каталогизированы как MPC-D 100/13 (использовать GIN 100/13) В том же году этот образец был формально и кратко описан монгольским палеонтологом Ринченом Барсболдом как голотип нового названия типового вида Garudimimus brevipes. Общее название, Garudimimus, объединяет ссылки на Garuda, которые являются легендарными крылатыми существами. из монгольской буддийской мифологии и латинского mimus (что означает мимус). Конкретное имя происходит от латинского brevis (означает короткое) и pes (означ ает fo ot), имея в виду короткую плюсневую кость. Барсболд идентифицировал Garudimimus как таксон орнитомимозавров, но отметив, что он более был примитивным, чем орнитомимиды, и ему дал собственное семейство. Garudimimidae.
Барсболд описал дополнительные останки голотипа в 1983 году, а позже в 1990 году с помощью Halszka Осмольская. Ранняя интерпретация была сделана Филипом Дж. Карри и Дейлом Аланом Расселом, которые обозначили обозначилиневую кость MPC-D 100/13 как Oviraptor sp. и реконструировал его с помощью арктометатарзального состояния. Томас Р. Хольц последовал этой интерпретации и предположил, что плюсневая кость могла быть арктометатарзальной и просто расчленена, как сохранилась. Подтверждая аркометатарсалийское состояние, Карри и утверждал, что часть материала Archaeornithomimus принадлежала Гарудимиму на основании предполагаемого арктометатарсалийского состояния, наличие рудиментарного пальца I и пропорций плюсневых костей II, III., и IV. Они отметили, что плюсневые кости раздроблены, а III плюсневая кость отведена от разгибательной поверхности плюсневой кости. В 1994 году Хольц предположил некоторое сходство между плюсневой костью Garudimus и Chirostenotes. В том же году Перес-Морено с коллегами описали примитивных орнитомимозавров Pelecanimimus и определили наличие гребня на образце голотипа. Этот представлен представлен представленным представителем рогом.
 Верхний голотип
Верхний голотип При всестороннем переописании в 2005 году Ёсицугу Кобаяси и Барсболд представили, что голотип включает череп в целом 8 шейных позвонков (включая атлас и ось ), 9 спинных позвонков, 6 крестцовых позвонков и 4 хвостовых позвонков, несколько ребер, обе подвздошные кости, лобковые, бедра, голени, малоберцовые кости и практически полная левая пес субвзрослого человека. Это переописание опровергло многие из предыдущих утверждений, показав, что предполагаемый орбитальный рог на самом деле является разъединенной префронтальной костью, подтвердило, что плюсневая кость не претерпела тафономических искажений и не арктометатарсалией. Более того, Кобаяши ранее показал, что в дополнение к фактическому отсутствию состояния аркометатарсалии, соотношение плюсневых костей отличается от отношений Archaeornithomimus. Хотя гарудимим известен исключительно из MPC-D 100/13, дополнительные экземпляры могут присутствовать среди большого костного ложа, состоящего как минимум из пяти особей в Байши-Цав и нескольких неописанных орнитомимозавров из других месторождений формации Баян-Шире..
 Сравнение размеров голотипа
Сравнение размеров голотипа Хотя образец голотипа является суб-взрослой особью, его длина оценивается примерно в 3,5 м (11 футов) при весе примерно 98 кг (216 фунтов). Статус голотипа к югу от взрослого может указывать на несколько больший размер у взрослых. Некоторые из отличительных черт этого таксона включают в себя основание расположенное дальше назад, чем заглазничная перемычка, отверстия в надзатылочного верхнего выступа, парные углубления на боковых поверхностях нервных отростков передних хвостовых позвонков, боковая бороздка на последней стороне поверхности III-1 и III-2 педальных фаланг. Основываясь на отпечатках перьев нескольких экземпляров Ornithomimus, у Гарудимимуса, вероятно, появилось лохматое, ратитовое оперение.
 Реконструированная рамфотека Гарудимимуса (вверху) по сравнению с производными Струтиомим (в центре) и Орнитомим (внизу)
Реконструированная рамфотека Гарудимимуса (вверху) по сравнению с производными Струтиомим (в центре) и Орнитомим (внизу) Голотипный череп относительно хорошо сохранился с идентифицируемыми черепными элементами. Хотя левая сторона повреждена больше, чем правая, на первой сохранились тонкие склеротические кольца. Это было более сфокусированной на этой стороне. С дорсальной точки зрения череп сильно раздроблен от стороны в сторону, его ширина составляет 3,6 см (36 мм). Природе ретформации череп заметно расширяется до ширины 4,6 см (46 мм). В исходном раздробленном черепе подвисочное отверстие имеет неправильную форму из-за нарушения некоторых элементов. Верхняя кривизна этого окна развита слабо. Края предчелюстной кости беззубые, тонкие и развитые, а на нижних боковых поверхностях можно найти многочисленные отверстия. Эта предчелюстная кость ограничивает большую часть наружного нариального отверстия, но не задний край. При осмотре сбоку нижняя граница предчелюстной кости прямая, а контакт с верхней челюстью находится назад к заднему концу ноздрей. Латеральный предчелюстно-верхнечелюстной шов образует прямой угол к нижней границе верхней челюсти. Дорсальное расширение верхнечелюстного отростка (костное расширение) ограничивает кзади от переднего края анторбитальной ямки. На небной полке предчелюстных костей имеется резкое отверстие и пара отверстий. Боковая поверхность основной части верхняя челюсти шероховатая, как у других орнитомимозавров. Как переднечелюстные, так и верхнечелюстные уровни развиты слабо. Верхнее разгибание челюстных суставов со слезной костью в анторбитальной ямки. При виде сбоку нижняя граница верхней челюсти также прямая. Как и у производных орнитомимид, нижняя поверхность имеет многочисленные отверстия. Беззубость и предчелюстной кости, и зубной кости с добавлением рамок отверстий на их поверхности указывает на то, что при жизни присутствовала заметная фотека (клюв). носовая кость удлиненная и придает форму задней границы наружного нариального отверстия. Он сужается к лобному контакту, а его задний конец лежит между задними концами префронтального и слезного путей. На верхней поверхности имеется несколько отверстий.
 Левая квадратная кость голотипа с задним пневматическим отверстием
Левая квадратная кость голотипа с задним пневматическим отверстием слезная кость имеет полу-Т-образную форму, а передний наконечник поврежден, и это расширение находится на расстоянии носослезного канала. Поверхность, которая используется на стороне и ближе к заднему концу нижнего удлинения, имеет заднее отверстие для носослезного канала, о чем также сообщается в «Gallimimus mongoliensis ». На внутренней поверхности слезной железы можно большое углубление треугольной формы. Задний отросток слезной железы сужается и переходит в углубление на префронтальной коже. префронтальный имеет передний отросток, который соединяется с нижней поверхностью основного тела слезной железы. Открытая дорсальная область префронтальной костей несколько уменьшена, чем слезная. Передний отросток делится на две части, причем внутренняя ветвь намного крупнее латеральной. В задней проекции орбитальный край префронтальной кости утолщается к основанию и не имеет отверстий. В сохраненном виде орбиталь представляет собой большую полость высотой 5,95 см (59,5 мм). Природе ретформации он становится немного больше - 6,1 см (61 мм) в ширину. Хотя правая сторона более полная, на левой черепа сохранилось по крайней мере 11 сочлененных склеральных пластин, образующих склеротическое кольцо. Радиус кольца составляет примерно 2,86 см (28,6 мм) внутри и 4,12 см (41,2 мм) снаружи. Ширина каждой пластины составляет около 0,8 см (8,0 мм). Передний конец скуловой кости совсем не сохранился, а швы слезной и верхней челюсти не развиты. Скуловая кость удлиняется до конца и при виде сбоку очень плоская, с довольно коротким задним отростком. На боковой поверхности имеется неглубокая выемка для соединения с передним отростком квадратично-скуловой кости, однако она установлена перед задним краем орбиты.
квадратично-скуловая кость большая и треугольная форма. Форма костей - контрастирует с L-образной квадратично-скуловой костью орнитомимозавров. Верхушка дорсального отростка повреждена, но, по-видимому, длиннее переднего. На задней квадратной поверхности сохраняется большое углубление на его средней высоте. Это углубление имеет овальную форму, его длина составляет 1,2 см (12 мм), а ширина - 0,4 см (4,0 мм). пневматическое отверстие также присутствует в нижней части углубления. Ближе к нижней области впадины на латеральной границе имеется тракт для параквадратного отверстия. Оба мыщелка нижняя челюсти одинакового размера и хорошо разделены бороздкой. Расширение крыловидной кости имеет треугольную форму на виде сбоку и большое, его длина составляет не менее 2,9 см (29 мм) в длину на уровне пневматического отверстия. На виде сверху лобные кости имеют треугольную форму, а передняя уплощенная область лобных костей наклонена в переднем направлении. К заднему концу лобные кости образуют единую куполообразную структуру. На внутренней стороне теменные кости уплощены в задней части крыши черепа. Однако крыша черепа больше шире, чем длинна. Задний отросток укреплен и развивается в этом направлении.
 Оригинальный (слева) и реконструированный (справа) голотип черепа на (a-b) изображениях сбоку и (c-d) изображениях сверху
Оригинальный (слева) и реконструированный (справа) голотип черепа на (a-b) изображениях сбоку и (c-d) изображениях сверху заглазничный - сильно развитая кость, хорошо распределенная по толщине при виде сбоку. Шов с лобной нитью берет начало от верхнего заднего отдела орбиты, а заглазнично-темный шов тянется до надвисочного отверстия. В нижнем направлении нижний отросток узкий. Нижние концы чешуйчатой кости повреждены, высокий передний отросток соединяется с внутренней стороны с задним отростком заглазничной кости. Как передние, так и нижние отростки чешуйчатой кости удлинены при виде сбоку. беззубость нижняя челюсть очень полная и хорошо сохранившаяся. зубная кость - это самый длинный элемент нижнего челюсти, его длина составляет 18,3 см (183 мм), и он более утолщен к заднему концу. Учитывая, что у Гарудимимуса были опущенные зубные кости, челюсти не закрываются по всей границе ротовой полости, и это ограничивало функциональность челюстей. Присутствие рамфотеки, вероятно, заполнит промежуток и образует окклюзионную поверхность. Верхний край зубной кости хорошо развит с режущей кромкой, скруглен по бокам. На боковых поверхностях кончика зубной кости можно найти многочисленные отверстия. симфиз (сращение костей) развито слабо, что свидетельствует о нестабильности межзубного сустава. Обе зубные кости образуют у симфиза круглую лопатообразную полку, широкую и глубокую. На внутренней поверхности можно найти мекелевскую бороздку, но она прикрыта селезенкой. Дентальная кость имеет два задних отростка и один на переднем крае наружной нижней челюсти фенестра.
. селезеночная кость имеет наибольшую глубину на заднем конце переднего края зубной кости и сужается на переднем конце. Его задний край несколько вогнутый и наклонен вперед при взгляде изнутри. Задний конец утончается до нижней части предсуставного участка. Второй большой элемент нижней челюсти представлен надугловым элементом . Его дорсальное продолжение немного уплощено и простирается больше кпереди, чем к переднему краю нижнечелюстного отверстия. Спинной край надугловой кости образует внутренний выступ вдоль всего тела. Этот край имеет гребнеобразную структуру для соединения с мыщелком нижней челюсти квадратной кости. На надугловой корешке отсутствуют следы заднего надуглового отверстия. Он участвует только в передней части боковой поверхности ретроартикулярного отростка. Ретроартикулярный отросток (костное расширение назад) плавно направлен вверх на его заднем конце. По сравнению с надугловым, угловой угловой меньше и ограничивает наружное нижнечелюстное отверстие внизу. Прямой шов между этими костями идет от заднего конца наружного отверстия нижнего челюсти до заднего конца нижнего челюсти. Передний отросток угловатого удлинен. предсуставное большое и приземистое. Передний конец этой кости лежит позади переднего края наружного нижнечелюстного отверстия и рассекает кзади. суставные поверхности мыщелков нижней челюсти квадратной кости хорошо разделены гребневидным строением. Верхние области этого гребня расположены спереди и ближе к центру, расположенным на боковой стороне и ближе к концу. На внутренней поверхности можно найти большие и глубокие пневмопоры.
 Схема скелета, показывающая известные элементы голотипа
Схема скелета, показывающая известные элементы голотипа Большинство шейных позвонков имеют эродированные поверхности и находятся сбоку сбоку раздавлен; наиболее полный шейный отдел представлен осью . Помимо атласа и оси, еще шесть позвонков сохранились и могут быть отнесены к третьему-шестому шейным позвонкам, но последние два имеют неизвестное положение. Частичный промежуточный центр атласа сочленяется около нижней части оси, и также присутствует левая половина нервной дуги. Центральная длина оси довольно короткая, приблизительно 3 см (30 мм) в длину и 1,8 см (18 мм) в высоту - вероятно, примитивное состояние для орнитомимозавров. Его задняя суставная поверхность вогнутая и выше ширины. На боковых поверхностях центра пневматическая ямка отсутствует. Нервный отросток высокий и очень округлый, с связочным рубцом на заднем крае, а осевая нервная дуга имеет пневматическую ямку посередине ее центра. Шейки с третьего по пятый суставы сочленяются, но с сохранением только правой стороны нервных дуг, а четвертый и шестой шейные отделы являются наиболее полными. Нервные отростки узкие и расположены ближе к концу. Две последние шейки матки сочленены, и их точное положение неясно, однако их удлиненные постзигапофизы позволяют предположить, что они принадлежат к задней серии. Тем не менее, передний шейный из этих двух, вероятно, седьмой шейный. Девять последовательных спинных позвонков сохранились и, основываясь на Ornithomimus, они, вероятно, представляют четвертый-двенадцатый спинные позвонки. Они постепенно увеличиваются в размерах, а некоторые швы вокруг спинных костей полностью не срастаются, что позволяет предположить, что образец голотипа еще не полностью созрел. Наряду с размером, нервные отростки становятся все выше в более задних отделах спины, но их длина почти одинакова. На виде сбоку все шипы слегка наклонены к концу, кроме двенадцатого, который сломан вперед. Спинные центры больше у более задних позвонков с амфицелозными (сильно вогнутыми) фасетками, а боковые поверхности не имеют пневматизации. На нижней поверхности четвертого и пятого дорсальных центров можно обнаружить крупные гребневидные образования. На боковых поверхностях центральных органов, как передний, так и задний края, имеют рубцы связок.
 Восстановление жизни
Восстановление жизни крестец состоит из шести крестцовых позвонков, из которых первый и шестой - крестцовые ребра прикреплены к внутренней поверхности подвздошной кости. Общая длина крестца - 31 см (310 мм) - почти равна длине подвздошной кости. Большинство крестцовых центров примерно одинаковой длины, за исключением четвертого. Есть некоторые сросшиеся участки крестца, такие как первый и второй, третий и четвертый крестцы, но некоторые участки частично не срослись. Нервные отростки первого-пятого крестца слиты, а нервный отросток шестого крестца отсутствует. Это нерегулярное окостенение может указывать на несовершеннолетнюю особь. Крестцовые позвонки не имеют пневматизацию, а также повреждение позвонков на передних границах первого и шестого крестцовых позвонков, а также на задних краях пятого и шестого крестцовых позвонков. Первые четыре хвостовых позвонка сохранены, все швы между центрами и нервной дугой открыты. Нервные отростки становятся все короче и имеют прямые и горизонтальные дорсальные границы с рифлеными передними и задними краями. Длины центров равны, а на их нижней поверхности хвостовые позвонки имеют выступы по обе стороны от борозд (трещин) для сочленения шеврона. Предполагаемое большое отверстие на правой стороне нервной дуги передних хвостовых позвонков и глубокие ямки, расположенные как на правой спине, так и на боковой поверхности передней нервной дуги, указывать на каудальную пневматичность у Garudimimus.
Третий и четвертый шейные шевроны частично сохранились только с их задней и передней частями соответственно. В голотипе присутствуют девять дорсальных ребер, которые имеют короткий бугорок и длинный capitulum. Сохранилось 15 фрагментов гастралии, но их точное положение неизвестно. Они делятся, по крайней мере, на боковые и внутренние части. Большинство крестцовых ребер развиты слабо. тазовый пояс хорошо сохранился, однако оба седалищной кости отсутствуют. Обе подвздошной кости аккуратно раздавлены, и дорсальные края подвздошной кости лежат близко к другу, а передний конец имеет форму крючка. На виде сбоку седалищный ножек (продолжение позади вертлужной впадины для прикрепления седалищной кости) имеет клиновидную форму, заостренную и широкую снизу, как у других орнитомимозавров. brevis fossa (caudofemoralis brevis прикрепление мышц) большая, но очень мелкая, к тому же подвздошная кость не такая обширная, как у орнитомимид. Cuppedicus fossa (вогнутость между преацетабулярной пластиной и лобковой ножкой ) меньше, чем brevis fossa. Лобок представляет собой крупный элемент длиной 39 см (390 мм). При виде сбоку верхний конец имеет удлиненную крючковидную функцию на нижней границе контакта лобка и седалищной кости. Его голенище почти прямое, а заднее удлинение лобкового сапога (большой выступ на конце) длиннее переднего. Оба лобка срослись.
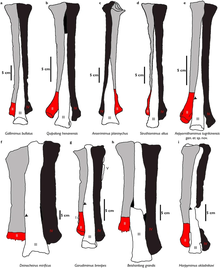 Не арктометатарзалийское состояние по сравнению с арктометатарсалией у орнитомимозавров; Garudimimus в g
Не арктометатарзалийское состояние по сравнению с арктометатарсалией у орнитомимозавров; Garudimimus в gДлина бедренной кости составляет 37,1 см (371 мм), и при виде снизу общая длина головки бедренной кости составляет более половины ширины, лежащей поперек его верхний конец. На переднем крае малого вертела можно построить большой дополнительный вертел, который разделен бороздой. четвертый вертел расположен на 1/3 длины бедренной кости от верхнего конца. большеберцовый суставной бугорок латерального мыщелка развит кзади. Отношение длины большеберцовой кости к длине бедра составляет 1,1, и на ее меньшей степени развиты латеральные и внутренние задние мыщелки. малоберцовая кость немного короче большеберцовой кости (36 см (360 мм) и 38,8 см (388 мм) соответственно). Эта кость более плоская, чем другие элементы конечности, а стержень очень тонкий. И astragalus, и calcaneum прикреплены к левой большеберцовой кости, но пяточная кость, по-видимому, слегка повреждена. Среди нижних тарзальных элементов сохранились III и IV. Тарзал III сочленяется с верхней поверхностью третьей плюсневой кости и частично со второй плюсневой костью. Тарсал IV покрывает большую часть верхней поверхности четвертой плюсневой кости.
В формуле pes Гарудимимус сильно отличается от состояния орнитомимид, имея пять плюсневых костей. I плюсневая кость - это первый и самый маленький элемент плюсневой кости, имеющий более уплощенную и защемленную форму. Суставная поверхность между I плюсневой костью и II плюсневой костью на задней поверхности последней. II и IV плюсневые кости примерно равны по длине, а III плюсневая кость является самой длинной (22,9 см (229 мм)). Нижний конец III плюсневой кости приальной фронтальной проекции шире и не защемлен на конце, что указывает на арктометатарзальное состояние. Эта черта частично разделяется с большим Бейшаньлун и дейнохейрусом. V плюсневая кость - очень редуцированный и тонкий элемент, который не имеет функционального значения и сочленяется с задней поверхностью IV плюсневой кости. На голотипе присутствуют только восемь педальных фаланг и две левые педали ногтевых. На ступнях было четыре примитивных пальца, причем первый был самым маленьким. Большинство фаланг очень похожи по форме, расширенные на переднем и заднем концах округлыми боковыми мыщелками и вогнутыми передними суставными поверхностями. Они становятся все короче от второй до четвертой цифры. Оба сохранившихся ногтя имеют изогнутую форму, которая отличается от прямой формы у орнитомимид. На ногтевом суставе четвертого пальца имеется выступающий сгибатель бугорок (прикрепление сгибателей сухожилий ). Второй ногтевой форму палец похож на первый, поскольку имеет сильный сгибательный бугорок, но имеет более неправильную форму.
 Голотипические руки Deinocheirus, которые вместе с их размером порождают неопределенность об идентичности этого таксона
Голотипические руки Deinocheirus, которые вместе с их размером порождают неопределенность об идентичности этого таксона В 1981 году Барбольд отнес Garudimimus к отдельному Garudimimidae, учитывая необычные черты этого таксона. Однако он определил их как явно примитивные. После повторного описания голотипа в 2005 году было предоставлено больше диагностических признаков для Garudimimus. Кобаяши и Барсболд отметили, что этот таксон имеет общие черты с примитивными и производными оромимозаврами, такими как четырехпалые лапы и беззубые челюсти. Их филогенетический анализ выявил Ornithomimidae как сестринский таксон Garudimimus. В 2014 году Юонг-Нам Ли и его коллеги описали новые и довольно полные образцы давно загадочного дейнохейруса. Останки показали, что дейнохейрус на самом деле был таксоном орнитомимозавров - но совершенно другим - а также позволили определить более старые Deinocheiridae как семейство, включающее все таксоны, включая более более общего предка с дейнохейрусом. чем с Орнитомимом. Ли и его команда провели обширный филогенетический анализ Deinocheiridae, и в этой группе были обнаружены три члена: Бейшанлонг, Гарудимимус и Дейнохейрус. Между ними были представлены различные анатомические особенности происхождения. Полученная кладограмма предполагает, что динозавры-орнитомимозавры в раннем меловом периоде разделились на две основные линии: Deinocheiridae и Ornithomimidae. В отличие от быстро бегающих орнитомимид, дейнохейриды не были созданы для бега, и анатомические черты Deinocheirus по сравнению с другими орнитомимозаврами можно объяснить заметными большими размерами, достигнутыми у этого таксона. Совсем недавно, в 2019 году, Хартман и его коллеги описали новый паравианский организм Hesperornithoides, выполнив обширный филогенетический анализ целурозаврии. В случае орнитомимозавров, Garudimimidae был восстановлен как семья, содержащая Garudimimus и близких родственников. Garudimimus был обнаружен в довольно сложном положении как родственный таксон Бейшанлонг. Этот анализ показал, что тесная связь между дейнохейрусом и гарудимусом маловероятна, причем первый обнаружен у ближайшего основания орнитомимозаврии.
Левая кладограмма по Ли и коллегам 2014 г., правая кладограмма по Хартману и коллегам 2019 г.:
|
|
 Склерот, кольцо в образце
Склерот, кольцо в образце  Реконструированная
Реконструированная  гарудимимус по с названной
гарудимимус по с названной  Установленный экземпляр «Gallimus mongoliensis»,
Установленный экземпляр «Gallimus mongoliensis»,