
Войти
| Овираптор Временной диапазон: поздний мел, ~75–71 млн лет PreꞒ Ꞓ О S D C п Т J K Стр. N ↓ | |
|---|---|
 | |
| Образец голотипа Овираптора | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Клэйд: | Динозаврия |
| Клэйд: | Заурисхия |
| Клэйд: | Тероподы |
| Надсемейство: | † Caenagnathoidea |
| Семья: | † Oviraptoridae |
| Род: | † Овираптор Осборн, 1924 г. |
| Типовой вид | |
| †Овираптор филоцератопс Осборн, 1924 г. | |
| Синонимы | |
| |
Овираптор (означает «яичный Seizer» или «яйцо вор») является родом из овирапторовидного динозавра, который жил в Азии в течение позднего мелового периода. Первые останки были собраны из свиты Djadokhta из Монголии в 1923 году во время палеонтологической экспедиции под руководством Роя Чепмена Эндрюса, и в следующем году в роде и типа вид овираптор philoceratops были названы Генри Fairfield Osborn. Название рода относится к первоначальной мысли о привычках кражи яиц, а конкретное название было предназначено для подкрепления этой точки зрения, указывающей на предпочтение цератопсовых яиц. Несмотря на то, что к роду были отнесены многочисленные экземпляры, Oviraptor известен только по одному частичному скелету, считающемуся голотипом, а также по гнезду из примерно пятнадцати яиц и нескольких небольших фрагментов молоди.
Овираптор был довольно маленьким пернатым овирапторидом, его длина оценивалась в 1,6 м (5,2 фута) при весе 33–40 кг (73–88 фунтов). У него была широкая нижняя челюсть с черепом, который, вероятно, имел гребень. И верхняя, и нижняя челюсти были беззубыми и имели роговой клюв, который использовался во время кормления вместе с крепкой морфологией нижних челюстей. Руки были хорошо развиты, удлинены, оканчивались тремя пальцами с загнутыми когтями. Как и другие oviraptorids, Овираптор были длинные задних конечностей, которые имели четыре носком ноги, с первым носком снижается. Хвост, вероятно, был не очень удлинен и заканчивался пигостилем, поддерживающим большие перья.
Первоначальные отношения Oviraptor были плохо изучены в то время и были отнесены к неродственным Ornithomimidae первоначальным описателем, Генри Осборном. Однако повторные исследования, проведенные Ринченом Барсболдом, доказали, что Oviraptor был достаточно отличным, чтобы основывать отдельное семейство, Oviraptoridae. При первом описании Овираптора интерпретировали как динозавра, который похитил яйца и поедал яйца, учитывая тесную связь голотипа с гнездом динозавров. Тем не менее, находки многочисленных овирапторозавров в позах гнездования показали, что этот экземпляр на самом деле вынашивал гнездо, а не крал и не питался яйцами. Кроме того, сообщалось об обнаружении останков маленького птенца или птенца, связанных с голотипом, что еще больше способствует заботе родителей.
 В Flaming Cliffs Монголии в пустыне Гоби. Останки голотипа Овираптора были найдены в этой высоко окаменелой местности джадохтинской свиты.
В Flaming Cliffs Монголии в пустыне Гоби. Останки голотипа Овираптора были найдены в этой высоко окаменелой местности джадохтинской свиты. Первые остатки овираптора были обнаружены на красноватых песчаниках в позднем меле формации Djadokhta из Монголии, в местности Баина ДЗАКА (также известную как Flaming Cliffs ), во время палеонтологической экспедиции в 1923 году экспедиция под руководством Североамериканского натуралист Рой Чепмен Эндрюса и закончилась открытием трех новых для науки ископаемых останков теропод, включая останки Овираптора. Они были официально описаны североамериканским палеонтологом Генри Фэрфилдом Осборном в 1924 году, который на основе нового материала назвал роды Oviraptor, Saurornithoides и Velociraptor. Конкретный род Oviraptor был создан с типовым видом O. philoceratops на основе голотипа AMNH 6517, частичной особи без задней части скелета, но с сильно раздробленным черепом, частичными шейными и спинными позвонками, грудными элементами, включая фуркулу с левой стороны. рука и часть кисти, левая подвздошная кость и некоторые ребра. Соответственно, этот экземпляр был найден лежащим над гнездом из примерно 15 яиц - гнездом, внесенным в каталог как AMNH 6508 - с черепом, отделенным от яиц всего на 10 см (100 мм) осадка. Учитывая тесную близость обоих образцов, Osborn интерпретированы овираптором как динозавр с яйцом питающихся привычек, и объяснил, что родовое название, Овираптор, является латинский для «яичного Seizer» или «яичного вора», из - за ассоциации окаменелостей. Специфическое название, филоцератопс, означает «любовь к яйцам цератопсов», что также является результатом первоначальной мысли о гнезде, относящемся к протоцератопсу или другому цератопсу. Однако Осборн предположил, что имя Овираптор могло отражать неправильное восприятие этого динозавра. Кроме того, Осборн обнаружил, что Овираптор похож на неродственных - в то время, однако, считавшихся родственными - быстроходных орнитомимидов, основанных на беззубых челюстях, и назначил Овираптора к Орнитомимидам. Осборн ранее сообщал о таксоне как «Fenestrosaurus philoceratops», но позже это было дискредитировано.
 Гнездо AMNH 6508, обнаруженное в ассоциации с голотипом.
Гнездо AMNH 6508, обнаруженное в ассоциации с голотипом. В 1976 году монгольский палеонтолог Ринчен Барсболд отметил некоторые несоответствия в таксономическом расположении Oviraptor и пришел к выводу, что этот таксон существенно отличается от орнитомимид по анатомическим признакам. Исходя из этого, он построил Oviraptoridae, чтобы содержать Oviraptor и близких родственников. После первоначального описания Овираптора Осборном гнездо яйца, связанное с голотипом, было принято как принадлежащее протоцератопсу, а овирапториды в значительной степени считались тероподами, питающимися яйцами. Тем не менее, в 1990-х годах открытие множества гнездящихся и гнездящихся экземпляров овирапторид доказало, что Осборн был прав в своем осторожном отношении к названию овираптора. Эти находки показали, что овирапториды насиживают и защищают свои гнезда, приседая на них. Эта новая линия свидетельств показала, что гнездо, связанное с голотипом Овираптора, принадлежало ему, и особь на самом деле вынашивала яйца в момент смерти, а не охотилась на них.
 Навесной MPC-D 100/42; образец, который стал основным эталоном для овираптора, но позже был опровергнут из-за черепных различий
Навесной MPC-D 100/42; образец, который стал основным эталоном для овираптора, но позже был опровергнут из-за черепных различий После присвоения Oviraptoridae названия в 1976 году Барсболд передал Oviraptor еще шесть экземпляров, в том числе два конкретных экземпляра под номерами MPC-D 100/20 и 100/21. В 1986 году Барсболд понял, что последние два не принадлежат к роду, а представляют нового овирапторида: Conchoraptor. Большинство других особей также вряд ли принадлежат самому овираптору, и они были отнесены к другим овирапторам. Частичная особь, также с яйцами из формации Баян Мандаху в Монголии, была упомянута в 1996 году Донг Чжимингом и Филипом Дж. Карри, образец IVPP V9608. Однако в 2010 году Николас Р. Лонгрич и два последних палеонтолога выразили свою неуверенность в отношении этого направления, поскольку существует несколько анатомических различий, таких как пропорции фаланг кисти. Они пришли к выводу, что этот экземпляр был другим и неопределенным видом, не относящимся к этому таксону. В 1981 году Барсболд передал образец MPC-D 100/42 Овираптору, очень хорошо сохранившейся и довольно полной особи из джадохтинской свиты. Поскольку известные элементы Oviraptor были настолько фрагментарными по сравнению с другими членами, MPC-D 100/42 стал основным эталоном / изображением этого таксона, получив заметное обозначение в научной литературе как Oviraptor philoceratops.
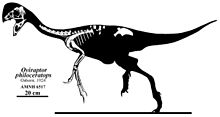 Скелетная реконструкция голотипа образца
Скелетная реконструкция голотипа образца Эта концепция была опровергнута Джеймсом М. Кларком и его коллегами в 2002 году, которые отметили, что этот экземпляр с высоким гребнем имеет больше общих черт черепа с Ситипати, чем с Овираптором, который фактически не сохраняет гребень, и может представлять второй вид прежнего или целый новый род. В 1986 году Барсболд описал второй вид овирапторов, «O. mongoliensis», на основе образца MPC-D 100 / 32a, который происходит из формации Немегт. Однако повторное исследование, проведенное Барсболдом в 1997 г., обнаружило достаточно различий в этом образце, чтобы назвать новый род Rinchenia, но он не описал его формально, и этот новый овирапторид остался как nomen dubium. Это было исправлено польским палеонтологом Хальской Осмольской и командой в 2004 году, официально назвав таксон Rinchenia mongoliensis. Североамериканский палеонтолог Марк А. Норелл и его коллеги в 2018 году сообщили новый экземпляр овираптора: AMNH 33092, который состоит из берцовой кости и два плюсны из птенца или очень небольших несовершеннолетних. AMNH 33092 был обнаружен вместе с голотипом и, вероятно, был частью гнезда. Овираптор теперь известен по голотипу с ассоциированными яйцами и по молоди / птенцу.
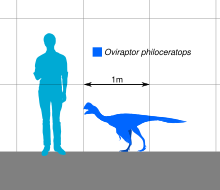 Сравнение размеров голотипа
Сравнение размеров голотипа Образец голотипа оценивается в длину 1,6 м (5,2 фута) и вес от 33 до 40 кг (от 73 до 88 фунтов). Хотя у голотипа в основном отсутствует задняя часть скелета, вполне вероятно, что у овираптора были две хорошо развитые задние конечности, которые заканчивались тремя функциональными пальцами, причем первый был рудиментарным, а также относительно уменьшенным хвостом. Как видно у родственных овирапторид, руки были покрыты удлиненными перьями, а хвост заканчивался пигостилем, который, как известно, поддерживает веер из перьев.
Череп из Овираптора был глубоким и укорочен с большим fenestrae (отверстия) по сравнению с другими динозаврами, и меры около 17,9 см (179 мм) длиной, как сохранились. Однако фактическая длина может быть больше, учитывая, что у голотипа черепа отсутствуют некоторые области, такие как предчелюстная кость. На голотипе черепа почти полностью отсутствует гребень, однако верхние поверхности сросшихся теменных и лобных костей указывают на то, что он, вероятно, имел хорошо развитый гребень, поддерживаемый носовыми и предчелюстными костями (в основном последними) рострума. У овираптора была удлиненная верхняя челюсть и зубная кость, что могло привести к более вытянутой морде по сравнению с сильно коренастыми челюстями других овирапторид. Вкус является жесткой, расширен ниже линии челюсти и образованные межчелюстных костей, сошников и челюстей. Как и у других овирапторид, у него могла быть пара зубчатых выступов на небе, направленных вниз. Как и у других овирапторид, ноздри (внешние ноздри) были относительно небольшими и располагались высоко на черепе. Овираптор был беззубые челюсти, которые заканчивались прочным, попугаи -как rhamphotheca (роговой клюв). Кривизна кончика зубной кости была понижена, но менее выражена, чем у других овирапторид, таких как Citipati. В целом нижняя челюсть представляет собой короткую и глубокую кость длиной 19,5 см (195 мм).
 Восстановление жизни
Восстановление жизни Как и у большинства овираптордов, нервные отростки голотипа шейных позвонков были короткими, а нервные дуги имели Х-образную форму. Однако в задних позвонках шипы становятся более выраженными. Зигапофизы первых шейных позвонков расположены параллельно друг другу, а постзигапофизы, по-видимому, существенно не расходятся от средней линии, в основном аналогично Citipati. В шейные ребра были присоединены к позвонков в голотипу. Нервные шипы имеют прямоугольную форму в переднем ряду спинных позвонков, если смотреть сбоку, и больше, чем шипы шейных позвонков. На переднем переднем спинном позвонке можно найти несколько плевроцелей (небольших отверстий с воздушным зазором), которые похожи на таковые у Хаана.
Вилка из Овираптор очень отличается от других oviraptorids в том, срединный киль на передней поверхности hypocleidium-а вниз направлены проекции в центре вилка. Кость V-образная, в поперечном сечении округлая, с удлиненным шиповидным гипоклеидием, межключичный угол около 90 °. Скапулокоракоид слит в голотипе, однако, коракоид сильно поврежден. Лопатка слегка поклонился и измеряет 23 см (230 мм) в длину. Овираптор имел относительно вытянутую руку, состоящую из плечевой кости, радиус, локтевой кости, и манус. Фаланговая формула овираптора была 2-3-4, как и у большинства других теропод и овирапторид. Рука овираптора имела три тощие и птичьи пальцы с каждым пальцем, завершившимся в стороны в сторону плоской и загнутые назад unguals ( коготь кость). В отличие от некоторых oviraptorids, Овираптор не пострадал сокращение второго и третьего пальца по отношению к первому.
Рассматриваемый молодой Oviraptor AMNH 33092 сохраняет материал задних конечностей, включающий правую большеберцовую кость с III и IV плюсневыми костью. Его большеберцовая кость имеет длину 5,87 см (58,7 мм), что свидетельствует о значительно меньших размерах, чем голотип. Гнездо AMNH 6508 хранит яйца элонгатоолитид, каждое из которых имеет длину 14 см (140 мм) (некоторые из них неполные). Тем не менее, существует вероятность того, что тафономическое дробление могло привести к их сжатию до 2 см (20 мм).
 Рисование линии голотипа черепа Осборном
Рисование линии голотипа черепа Осборном Овираптор был первоначально связан с орнитомимидами Осборном из-за его беззубого клюва. Осборн также обнаружил сходство с хиростенотом, который до сих пор считается близким родственником овираптора. В 1976 году Barsbold возведено новое семейство содержит овираптором и его близких родственников, что делает Овираптор в тип рода по Oviraptoridae. Во время повторного описания голотипа черепа в 2002 году Кларком и его коллегами они отметили, что у овираптора была относительно удлиненная верхняя челюсть и зубная кость. Эти признаки менее выражены у производных овирапторид и позволяют предположить, что Oviraptor принадлежит к ближайшему основанию Oviraptoridae.
Приведенная ниже кладограмма следует за анализом, проведенным Грегори Фанстоном и его коллегами в 2020 году:
| Oviraptoridae |
| ||||||||||||||||||||||||
 Восстановлен голотип череп из овираптора
Восстановлен голотип череп из овираптора При первом описании Овираптора в 1924 году Осборном первоначально предполагалось, что он имел яйцеклетку - организм, питающийся яичной диетой - образ жизни, основанный на ассоциации голотипа с гнездом, которое, как считалось, принадлежало протоцератопсу. В 1977 году Барсболд выдвинул гипотезу сокрушительной челюсти. Он утверждал, что сила крепких нижних челюстей и, вероятно, рамфотеки (рогового клюва) была достаточно сильной, чтобы сломать раковины моллюсков, таких как моллюски, которые встречаются в той же геологической формации, что и Овираптор. Эти кости образуют часть основной кости верхней челюсти или верхней челюсти, которые сходятся посередине, образуя пару зубцов. Рамфотека и нижняя челюсть вместе с продолжением нескольких костей из неба могли быть инструментом для пирсинга. Барсболд также предположил, что овирапториды могли вести полуводный образ жизни, основанный на диете на основе моллюсков, высоком расположении носовых полостей, увеличенной мускулатуре хвоста и большем размере первого пальца руки. В тезисе конференции 1990 года Дэвид К. Смит представил остеологическую переоценку Овираптора, в которой он отверг утверждения Барсболда. Он не обнаружил никаких доказательств того, что передняя конечность специализировалась на движении в воде, а челюсти, вместо того, чтобы сохранять механизм дробления, сохраняют режущие поверхности. Поскольку череп беззубый, легко сложенный и не имеет нескольких сильных областей прикрепления мышц, Смит предположил, что листья могли быть важной частью диеты овираптора. Однако в 1995 году Норелл и его коллеги сообщили о фрагментированных останках ящерицы в полости тела экземпляра голотипа, предполагая, что Овираптор был частично плотоядным.
 Сравнение нижней челюсти нескольких родов овирапторозавров
Сравнение нижней челюсти нескольких родов овирапторозавров В 2008 году Стиг Олав К. Янсен сравнил черепа нескольких видов овирапторид с черепами птиц и черепах, чтобы выяснить, по каким свойствам можно предсказать наличие рамфотеки. Он обнаружил, что нижние челюсти овирапторид очень похожи на челюсти попугаев, а верхние челюсти больше похожи на челюсти черепах. Основываясь на этих наблюдениях, Янсен предположил, что овирапториды были всеядными, так как резко развитая рамфотека вместе с выступающими передними конечностями могла быть приспособлена для ловли и отрывания мелкой добычи. Более того, заостренные выступы неба помогли удерживать добычу. Янсен указал, что полностью растительноядная диета овирапторид кажется маловероятной, поскольку у них не хватало плоских и широких томий (режущих краев нижних челюстей) для жевания, и они не могли двигать нижними челюстями вбок. Однако он считал нижнюю челюсть достаточно сильной, чтобы в ней могли быть по крайней мере раздавленные элементы, такие как яйца, орехи или другие твердые семена.
Лонгрич и его коллеги в 2010 году также отвергли гипотезу о дурофагах (животных, которые практикуют дробление панцирей ), учитывая, что у таких животных обычно появляются зубы с широкой дробящей поверхностью. Заостренная форма зубных костей нижней челюсти предполагает, что у овирапторид вместо этого для стрижки пищи использовалась остроконечная рамфотека. Symphyseal (костный союз) область в передней части зубной кости, возможно, дал некоторую способность для дробления, но так как это был относительно небольшой площади, это, вероятно, не является основной функцией челюстей. Еще один аргумент против того, что они поедали моллюсков, заключается в том, что большинство овирапторид было обнаружено в отложениях, которые интерпретируются как представляющие в основном засушливые или полузасушливые среды, такие как овираптор в формации Джадохта. Команда также обнаружила, что овирапториды и дицинодонты имеют общие черты черепа, такие как короткие, глубокие и беззубые нижние челюсти; удлиненные зубные симфизы; удлиненные нижнечелюстные отверстия; и острое небо. Современные животные с челюстями, напоминающими челюсти овирапторид, включают попугаев и черепах ; последняя группа также имеет зубчатые выступы на небной области. Лонгрич и его коллеги пришли к выводу, что из-за сходства между овирапторидами и травоядными животными, основная часть их рациона, скорее всего, была сформирована из растительного материала. Челюсти овирапторид, возможно, были специализированы для обработки пищи, такой как ксерофитная растительность - растительность, адаптированная для окружающей среды с небольшим количеством воды - которая могла бы расти в их засушливой среде, но это невозможно продемонстрировать, так как мало что известно о палеофлора пустыни Гоби.
Однако в 2018 году Фунстон и его коллеги поддержали гипотезу сокрушительной челюсти. Они отметили, что коренастый рострум и крепкая нижняя челюсть овирапторид на самом деле предполагают сильный и кусающий укус, который очень похож на укус попугаев. Фунстон и его коллеги считали, что эти анатомические особенности овирапторид соответствуют плодоядной диете, включающей орехи и семена.
 Гнездящийся экземпляр ситипати
Гнездящийся экземпляр ситипати  Гнездо овираптора AMNH 6508
Гнездо овираптора AMNH 6508 С момента описания эмбрионального экземпляра Citipati в 1994 году овирапториды стали более понятными: вместо того, чтобы питаться яйцами, они фактически вынашивали гнезда и заботились о них. Этот образец показал, что голотип Овираптора, вероятно, был половозрелой особью, которая погибла, насиживая связанное с ним гнездо с яйцами. Это новое поведение овирапторид стало более ясным после отчета и краткого описания взрослого экземпляра Citipati, гнездящегося в 1995 году, Норелла и его коллег. Экземпляр был найден на вершине кладок для яиц, его задние конечности согнуты симметрично с каждой стороны гнезда, а передние конечности покрывали периметр гнезда. Эта задумчивая поза встречается сегодня только у современных птичьих динозавров и поддерживает поведенческую связь между последней группой и нептичьими динозаврами. В 1996 году Донг и Карри описали новую гнездящуюся особь овирапторид из формации Баян Мандаху. Он был найден лежащим на вершине гнезда, состоящего примерно из 6 сохранившихся яиц, которые были заложены в виде холмика с круговым рисунком. Поскольку образец был найден над гнездом, его передние конечности покрывали яйца, а частично сохранившаяся задняя лапа - около центра гнезда, Донг и Карри предположили, что он был пойман и захоронен песчаной бурей во время инкубации. Они исключили возможность того, что овирапториды могут похитить яйца, поскольку они либо съели, либо инстинктивно покинули гнездо задолго до того, как оно было похоронено из-за песчаной бури или другого метеорологического явления.
В 1999 году Кларк и его команда подробно описали ранее описанный образец гнездования Citipati и кратко обсудили образец голотипа Oviraptor и его связь с гнездом AMNH 6508. Они отметили, что точное положение голотипа над гнездом неясно, поскольку они были разделены во время подготовки, и гнездо, по-видимому, не полностью завершено, сохранилось около 15 яиц, два из которых повреждены. Более того, полукруглое расположение гнезда указывает на то, что яйца откладывались попарно и по крайней мере в три кольца, причем изначально это гнездо было круглым, похожим на холмик.
Томас П. Хопп и Марк Дж. Орсен в 2004 году проанализировали насиживающее поведение вымерших и современных видов динозавров, включая овирапторид, чтобы оценить причину удлинения и развития крыльев и рулевых перьев. Учитывая, что наиболее полный экземпляр гнездящихся овирапторид - в то время гнездящийся экземпляр Citipati 1995 года - был обнаружен в позе, очень напоминающей птичью, с передними конечностями в почти сложенном положении, а грудная область, живот и ступни соприкасались с ними. яйца, Хопп и Орсен указали, что длинные перистые перья и перьевой покров, скорее всего, присутствовали в жизни. «Крылья» и хвост овирапторид обеспечили бы защиту яиц и детенышей от климатических факторов, таких как солнечный свет, ветер и осадки. Однако руки этого экземпляра не были чрезвычайно сложены, как у некоторых современных птиц, вместо этого они были более вытянутыми, напоминая стиль крупных нелетающих птиц, таких как страус. Вытянутое положение руки также похоже на поведение этой птицы, которая, как известно, гнездится большими кладками, такими как овирапториды. Основываясь на положении передних конечностей у гнездящихся овирапторид, Хопп и Орсен предположили, что выведение потомства является наследственной причиной удлинения крыльев и хвостовых перьев, поскольку существует большая потребность в обеспечении оптимальной защиты яиц и молоди.
 Восстановление жизни гнездящейся Немегтомайи. Овирапториды могли сидеть на своих гнездах во время инкубации.
Восстановление жизни гнездящейся Немегтомайи. Овирапториды могли сидеть на своих гнездах во время инкубации. В 2005 году Тамаки Сато и его команда сообщили о необычном образце овирапторид из формации Наньсюн. Этот новый образец, как было обнаружено, сохранил в основном область таза с двумя яйцами внутри и, таким образом, указывал на самку. Размер и положение яиц позволяют предположить, что овирапториды сохранили два функциональных яйцевода, но сократили количество овулировавших яиц до одного на яйцевод. Дэвид Дж. Варриккио и его коллеги в 2008 году обнаружили, что относительно большой размер кладок яиц у овирапторид и троодонтид больше всего похож на таковые у современных птиц, практикующих полигамное спаривание и обширную родительскую заботу самцов, таких как птицы- ратиты, предполагающие аналогичные привычки. Эта репродуктивная система, скорее всего, представляет собой наследственное состояние современных птиц, а забота о двух родителях (в которой участвуют оба родителя) является более поздним развитием. В 2014 году У. Скотт Персонс и его коллеги предположили, что овирапторозавры вторично нелетают, и некоторые черты их хвостов могут указывать на склонность к демонстративному поведению, например, демонстрацию ухаживания. Хвост некоторых овирапторозавров и овирапторид заканчивался пигостилем - костной структурой на конце хвоста, которая, по крайней мере, у современных птиц, используется для поддержки веера из перьев. Кроме того, хвост был особенно мускулистым и обладал ярко выраженной гибкостью, что, возможно, помогало при ухаживании.
 Завершите кладку яиц овирапторид из формации Наньсюн.
Завершите кладку яиц овирапторид из формации Наньсюн. В 2018 году Цзы-Руэй Ян и его коллеги определили слои кутикулы на нескольких яичных оболочках динозавров манирапторанов, в том числе овирапторид. Эти конкретные слои состоят из белков, полисахаридов и пигментов, но в основном из липидов и гидроксиапатита. У современных птиц они служат для защиты яиц от обезвоживания и вторжения микроорганизмов. Поскольку большинство экземпляров овирапторид были обнаружены в образованиях седиментации на основе калише, Ян и его коллеги предположили, что яйца, покрытые кутикулой, были бы репродуктивной стратегией, адаптированной для повышения их успешности вылупления в таком засушливом климате и окружающей среде.
В 2019 году Ян и его коллеги переоценили гипотезу терморегуляторной контактной инкубации с использованием полных гнезд овирапторид из формации Наньсюн и предоставили подробную реконструкцию архитектуры кладки овирапторид. Они отметили, что взрослые особи овирапторид в связи с гнездом не обязательно инкубировали яйца, поскольку они могли представлять самку в процессе откладки яиц, а многокольцевое сцепление предотвращало достаточную передачу тепла от родителя к внутренним кольцам яиц. Среднее гнездо овирапторид было построено как пологий холм с высокоорганизованной архитектурой: яйца, вероятно, были пигментированы и располагались парами, каждая пара располагалась в виде трех-четырех эллиптических колец. Поскольку родитель, вероятно, действовал из центра гнезда, в этой области не было яиц. Янг и его коллеги пришли к выводу, что стиль гнездования овирапторид настолько уникален, что им не хватает современных аналогов, поэтому использование репродукции овирапторид может быть не лучшим примером для информирования об эволюции репродуктивных стратегий птиц. Однако команде не удалось определить, вылупился ли молодой овираптор AMNH 33092 из гнезда, связанного с голотипом.
 Реставрация покоящегося овираптора джадохтинской свиты
Реставрация покоящегося овираптора джадохтинской свиты Овираптор известен из местонахождения Байн-Дзак в формации Джадохта в Монголии, которая восходит к позднему меловому периоду примерно от 71 до 75 миллионов лет назад. Палеосреда джадохтинской свиты интерпретируется как имеющая полузасушливый климат с песчаными дюнами и аллювиальными условиями, подобными современной пустыне Гоби. Полузасушливый степной ландшафт осушался прерывистыми ручьями и иногда подвергался воздействию пыли и песчаных бурь, а влажность была сезонной. Хотя считается, что это образование в значительной степени относится к хорошо сохранившимся высокозасушливым средам, несколько недолговечных водоемов были зарегистрированы в местонахождении Ухаа Толгод на основе речных отложений. Более того, считается, что позже, в кампанскую эпоху и в маастрихтский период, климат сместится к более влажной речной среде, наблюдаемой в формации Немегт.
Джадохтинская свита разделяется на нижнюю байн-дзакскую пачку и верхнюю тургругинскую пачку. Известные останки Овираптора были произведены членом Байн Дзак, который также дал динозавров Байноцератопс, Пинакозавр, Протоцератопс, Заурорнитоидес, Велоцираптор и Хальшкараптор. Другие динозавр фауна этого пользователя включает в себя, что в местности Ухаа Толгоды, состоящей из Apsaravis, Byronosaurus, читипатьте, Gobipteryx, Хаано, Колет, Shuuvuia, Tsaagan и Minotaurasaurus.
Поза голотипа Овираптора вместе с ассоциацией яиц позволяет предположить, что он оказался в ловушке над гнездом во время песчаной бури, а захоронение было относительно быстрым, учитывая, что у тела не было возможности полностью разъединиться или быть уничтоженным хищниками. Палеонтолог Кеннет Карпентер также согласился с тем, что песчаные бури, возможно, были наиболее вероятным случаем захоронения яиц, найденных в отложениях. Среди элементов череп стал особенно уплощенным и деформированным в процессе окаменения.