
Войти
 Усиление помогает видоизменению путем выбора против гибридов при вторичном контакте двух отдельных популяций вида.
Усиление помогает видоизменению путем выбора против гибридов при вторичном контакте двух отдельных популяций вида. Усиление - это процесс видообразования, где естественный отбор увеличивает репродуктивная изоляция (далее разделенная на презиготическая изоляция и постзиготическая изоляция ) между двумя популяциями видов. Это происходит в результате отбора, действующего против производства гибридных особей с низкой приспособленностью. Идея была первоначально разработана Альфредом Расселом Уоллесом и иногда упоминается как эффект Уоллеса . Современная концепция армирования берет свое начало от Феодосия Добжанского. Он предвидел вид, разделенный аллопатрически, где во время вторичного контакта две популяции спариваются, давая гибриды с более низкой приспособленностью. Естественный отбор является результатом неспособности гибрида производить жизнеспособное потомство; таким образом, представители одного вида, не спаривающиеся с представителями другого, имеют больший репродуктивный успех. Это способствует эволюции большей презиготической изоляции (различия в поведении или биологии, которые препятствуют образованию гибридных зигот). Усиление - один из немногих случаев, когда отбор может способствовать увеличению презиготической изоляции, напрямую влияя на процесс видообразования. Этот аспект был особенно привлекателен среди биологов-эволюционистов.
Поддержка подкрепления колебалась с момента ее появления, а терминологическая путаница и различия в использовании на протяжении истории привели к множеству значений и осложнений. биологи-эволюционисты выдвинули различные возражения относительно правдоподобности его возникновения. С 1990-х годов данные теории, экспериментов и природы преодолели многие из прошлых возражений, сделав подкрепление широко принятым, хотя его распространенность в природе остается неизвестной.
Было разработано множество моделей, чтобы понять его действие в природе., большинство из которых зависит от нескольких аспектов: генетики, популяционных структур, влияния отбора и брачного поведения. Эмпирическая поддержка армирования существует как в лаборатории, так и в природе. Задокументированные примеры обнаружены у широкого круга организмов: как позвоночных, так и беспозвоночных, грибов и растений. Вторичный контакт первоначально разделенных зарождающихся видов (начальная стадия видообразования) увеличивается из-за деятельности человека, такой как интродукция инвазивных видов или изменение естественных местообитаний. Это имеет значение для показателей биоразнообразия и может стать более актуальным в будущем.
У подкрепления сложная история: его популярность среди ученых со временем изменилась. Джерри Койн и Х. Аллен Орр утверждает, что теория подкрепления прошла три фазы исторического развития:
 Альфред Рассел Уоллес в 1889 году предположил, что изоляция может быть усилена формой отбора.
Альфред Рассел Уоллес в 1889 году предположил, что изоляция может быть усилена формой отбора. Иногда это называется эффектом Уоллеса, усиление было первоначально предложено Альфредом Расселом Уоллесом в 1889 году. Его гипотеза заметно отличалась от современной концепции тем, что она фокусировалась на постзиготической изоляции, усиленной групповым отбором. Феодосий Добжанский был первым представил подробное описание процесса в 1937 году, хотя сам термин не был введен до 1955 года У. Фрэнк Блэр. В 1930 году Рональд Фишер изложил первое генетическое описание процесса подкрепления в Генетической теории естественного отбора, а в 1965 и 1970 годах были проведены первые компьютерные симуляции для проверки его правдоподобие. Позже были проведены популяционно-генетические и количественные генетические исследования, показавшие, что совершенно непригодные гибриды однозначно приводят к усилению презиготической изоляции.
Идея Добжанского получила значительную поддержку; он предположил, что это иллюстрирует заключительный этап видообразования, например, после того, как аллопатрическая популяция вступает во вторичный контакт. В 1980-х годах многие биологи-эволюционисты начали сомневаться в правдоподобности этой идеи, основываясь не на эмпирических данных, а в значительной степени на росте теории, которая считала это маловероятным механизмом репродуктивной изоляции. В то время возник ряд теоретических возражений, которые рассматриваются в разделе «Аргументы против подкрепления» ниже.
К началу 1990-х годов популярность подкрепления среди биологов-эволюционистов возродилась; в первую очередь из-за внезапного увеличения количества данных - эмпирических данных исследований в лабораториях и, в основном, примеров, найденных в природе. Кроме того, компьютерное моделирование генетики и моделей миграции популяций обнаружило «нечто похожее на подкрепление». Самая последняя теоретическая работа по видообразованию была основана на нескольких исследованиях (особенно от Лиу и Прайса, Келли и Нура и Киркпатрика и) с использованием очень сложных компьютерных симуляций; все они пришли к схожим выводам: что подкрепление правдоподобно при нескольких условиях и во многих случаях проще, чем считалось ранее.
Существует путаница вокруг значения термина «подкрепление». Впервые он был использован для описания наблюдаемых различий в брачных криках у лягушек Gastrophryne внутри гибридной зоны вторичного контакта. Термин вторичный контакт также использовался для описания подкрепления в контексте аллопатрически разделенной популяции, испытывающей контакт после потери географического барьера. Эффект Уоллеса похож на армирование, но используется редко. разграничивает неполную постзиготическую изоляцию от полной изоляции, ссылаясь на неполную изоляцию как на подкрепление, а полностью изолированные популяции как на испытывающие смещение репродуктивного характера. Дэниел Дж. Ховард считал смещение репродуктивного характера либо ассортивным спариванием, либо дивергенцией признаков для распознавания партнера (особенно между симпатрическими популяциями). Подкрепление, по его определению, включало презиготическую дивергенцию и полную постзиготическую изоляцию. Servedio и Noor включают любое обнаруженное увеличение презиготической изоляции в качестве подкрепления, если это является ответом на отбор против спаривания между двумя разными видами. Койн и Орр утверждают, что «истинное подкрепление ограничено случаями, когда изоляция усиливается между таксонами, которые все еще могут обмениваться генами».
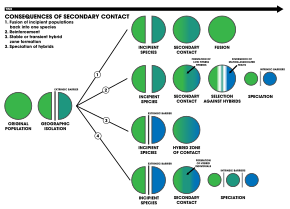 Четыре результата вторичного контакта:. 1. Внешний барьер разделяет популяцию вида на две части, но они вступают в контакт до того, как репродуктивная изоляция становится достаточной, чтобы привести к видообразованию. Две популяции сливаются в один вид. 2. Видообразование по армированию. 3. Две отдельные популяции остаются генетически разными, в то время как гибридные рои формируются в зоне контакта. 4. Геномная рекомбинация приводит к видообразованию двух популяций с дополнительным гибридным видом. Все три вида разделены внутренними репродуктивными барьерами
Четыре результата вторичного контакта:. 1. Внешний барьер разделяет популяцию вида на две части, но они вступают в контакт до того, как репродуктивная изоляция становится достаточной, чтобы привести к видообразованию. Две популяции сливаются в один вид. 2. Видообразование по армированию. 3. Две отдельные популяции остаются генетически разными, в то время как гибридные рои формируются в зоне контакта. 4. Геномная рекомбинация приводит к видообразованию двух популяций с дополнительным гибридным видом. Все три вида разделены внутренними репродуктивными барьерами Одной из самых сильных форм репродуктивной изоляции в природе является сексуальная изоляция: признаки у организмов, вовлекающие спаривание. Этот паттерн привел к мысли, что, поскольку отбор так сильно влияет на черты спаривания, он может быть вовлечен в процесс видообразования. Этот процесс видообразования под влиянием естественного отбора является подкреплением и может происходить при любом способе видообразования (например, географических способах видообразования или экологическом видообразовании ). Для этого необходимы две силы эволюции, которые действуют на выбор партнера : естественный отбор и поток генов. Отбор выступает в качестве основного двигателя подкрепления, поскольку он отбирает гибрид генотипов, которые имеют низкую- приспособленность, независимо от того, не влияют ли индивидуальные предпочтения на выживание и воспроизводство. Поток генов действует как основная сила противодействия подкреплению, поскольку обмен генами между особями, приводящий к гибридам, приводит к гомогенизации генотипов..
Бутлин изложил четыре основных критерия подкрепления, которые должны быть обнаружены в естественных или лабораторных популяциях:
После того, как происходит видообразование путем подкрепления, происходят изменения после полной репродуктивной изоляции (и последующей изоляции) форма репродуктивного смещения характера. Обычный признак появления подкрепления в природе - это смещение репродуктивного характера ; характеристики населения расходятся по симпатрии, но не по аллопатрии. Одна из трудностей в обнаружении состоит в том, что смещение экологического символа может привести к тем же самым образцам. Кроме того, поток генов может уменьшить изоляцию симпатрических популяций. Два важных фактора в исходе процесса зависят от: 1) конкретных механизмов, вызывающих презиготную изоляцию, и 2) количества аллелей, измененных мутациями, влияющими на выбор партнера.
В случаях перипатрического видообразования., подкрепление вряд ли завершит видообразование в случае, если периферически изолированная популяция вступает во вторичный контакт с основной популяцией. В симпатрическом видообразовании требуется отбор против гибридов; поэтому подкрепление может сыграть свою роль, учитывая эволюцию некоторых форм компромиссов в пригодности. В симпатрии часто наблюдаются модели сильной дискриминации при спаривании, которые приписываются подкреплению. Считается, что подкрепление является агентом игровой изоляции.
Основную генетику подкрепления можно понять с помощью идеальной модели двух гаплоидных популяций, испытывающих увеличение нарушение равновесия по сцеплению. Здесь отбор отклоняет комбинации с низкой пригодностью 











Существует альтернативная модель для устранения антагонизма рекомбинации, поскольку это может уменьшить ассоциацию между аллелями, которые связаны с приспособленностью, и ассортивными аллелями спаривания, которые не связаны. Генетические модели часто различаются по количеству признаков, связанных с локусами ; некоторые полагаются на один локус на каждый признак, а другие - на полигенные признаки.
Структура и модели миграции популяции могут влиять на процесс видообразования путем подкрепления. Было показано, что это происходит в модели острова, в которой существуют условия с нечастыми миграциями, происходящими в одном направлении, и в моделях симметричной миграции, когда виды равномерно мигрируют туда и обратно между популяциями.
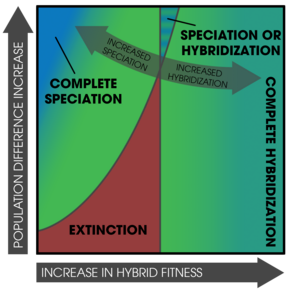 Пространство параметров, представляющее условия, в которых видообразование путем подкрепления может возникнуть. Здесь могут возникнуть три исхода: 1) исчезновение одной из исходных популяций; 2) исходные популяции могут гибридизоваться; 3) начальные популяции могут видоизменяться. Результаты определяются как исходной дивергенцией, так и уровнем приспособленности гибридов.
Пространство параметров, представляющее условия, в которых видообразование путем подкрепления может возникнуть. Здесь могут возникнуть три исхода: 1) исчезновение одной из исходных популяций; 2) исходные популяции могут гибридизоваться; 3) начальные популяции могут видоизменяться. Результаты определяются как исходной дивергенцией, так и уровнем приспособленности гибридов. Усиление может также происходить в отдельных популяциях, мозаичных гибридных зонах (неоднородное распределение родительских форм и субпопуляций) и в парапатрические популяции с узкими контактными зонами.
Плотность населения - важный фактор в подкреплении, часто в сочетании с вымиранием. Возможно, что при вторичном контакте двух видов одна популяция вымрет - в первую очередь из-за низкой гибкости приспособленности, сопровождающейся высокими темпами роста популяции. Исчезновение менее вероятно, если гибриды будут необратимыми вместо бесплодными, поскольку фертильные особи все еще могут выжить достаточно долго, чтобы размножаться.
Видообразование путем подкрепления напрямую зависит от отбора, способствующего усилению презиготической изоляции, и природа роли отбора в подкреплении широко обсуждалась с моделями, применяющими различные подходы. Селекция, действующая на гибриды, может происходить несколькими способами. Все производимые гибриды могут иметь одинаковую низкую пригодность, что дает большой недостаток. В других случаях отбор может отдавать предпочтение множественным и различным фенотипам, например, в случае мозаичной гибридной зоны. Естественный отбор может воздействовать на конкретные аллели как прямо, так и косвенно. При прямом отборе частота выбранного аллеля максимально благоприятна. В случаях, когда аллель выбирается косвенно, его частота увеличивается из-за другого связанного аллеля, подвергающегося отбору (неравновесное сцепление ).
Состояние гибридов при отборе может играть роль в постзиготической изоляции, поскольку Нестабильность гибридов (гибрид, неспособный созреть до пригодной взрослой особи) и бесплодие (неспособность полностью произвести потомство) запрещают поток генов между популяциями. Отбор против гибридов может даже быть вызван любой неудачей в получении партнера, поскольку он практически неотличима от бесплодия - каждое обстоятельство не приводит к появлению потомства.
Для того, чтобы произошло подкрепление, должно присутствовать некоторое начальное расхождение в предпочтениях партнера. Любые черты, способствующие изоляции, могут быть подвергаются подкреплению, например, сигналы спаривания (например, отображение ухаживания ), ответы сигнал, местоположение нерестилищ, время спаривания (например, сезонный разведение ), или даже яйцо восприимчивость. Люди могут также дискриминировать партнеров, которые различаются по различным признакам, таким как брачный зов или морфология. Многие из этих примеров описаны ниже.
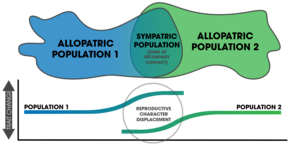 Две аллопатрические популяции вступают во вторичный контакт. В симпатрии дивергенция проявляется в изменении брачных черт. Эти паттерны смещения репродуктивных признаков, обнаруженные в популяциях видов, которые существуют в зонах перекрытия, указывают на то, что произошел процесс видообразования путем подкрепления.
Две аллопатрические популяции вступают во вторичный контакт. В симпатрии дивергенция проявляется в изменении брачных черт. Эти паттерны смещения репродуктивных признаков, обнаруженные в популяциях видов, которые существуют в зонах перекрытия, указывают на то, что произошел процесс видообразования путем подкрепления. свидетельство подкрепления происходит из наблюдений в природе, сравнительных исследований и лабораторные эксперименты.
Можно показать, что подкрепление происходит (или происходило в прошлом), путем измерения силы презиготической изоляции в симпатрической популяции по сравнению с аллопатрической популяцией того же вида. Сравнительные исследования этого позволяют определить крупномасштабные закономерности в природе для различных таксонов. Образцы спаривания в гибридных зонах также можно использовать для обнаружения подкрепления. Смещение репродуктивного характера рассматривается как результат подкрепления, поэтому во многих случаях в природе этот образец выражается в симпатрии. Повсеместность подкрепления неизвестна, но паттерны смещения репродуктивных признаков обнаруживаются во многих таксонах и считаются обычным явлением в природе. Изучение подкрепления в природе часто оказывается трудным, поскольку можно утверждать альтернативные объяснения обнаруженных закономерностей. Тем не менее, существуют эмпирические данные, подтверждающие наличие подкрепления в различных таксонах, и его роль в ускорении видообразования неоспорима.
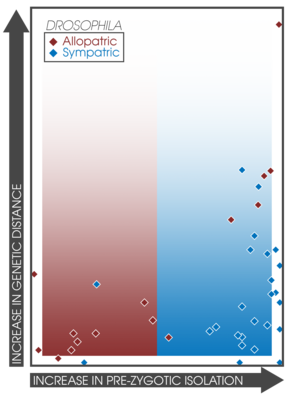 Презиготическая изоляция в парах аллопатрических (красный) и симпатрических (синий) видов Drosophila. Градиенты указывают на прогнозы подкрепления для аллопатрических и симпатрических популяций.
Презиготическая изоляция в парах аллопатрических (красный) и симпатрических (синий) видов Drosophila. Градиенты указывают на прогнозы подкрепления для аллопатрических и симпатрических популяций. Ожидается, что вспомогательное спаривание будет увеличиваться среди симпатрических популяций, испытывающих подкрепление. Этот факт позволяет напрямую сравнивать силу презиготической изоляции в симпатрии и аллопатрии между различными экспериментами и исследованиями. Койн и Орр исследовали 171 пару видов, собирая данные об их географическом характере, генетическом расстоянии и силе презиготической и постзиготической изоляции; обнаружение, что презиготическая изоляция была значительно сильнее в симпатрических парах, коррелируя с возрастом вида. Кроме того, сила постзиготической изоляции не различалась между симпатрическими и аллопатрическими парами. Это открытие подтверждает предсказания видообразования путем подкрепления и хорошо коррелирует с более поздним исследованием, в котором было обнаружено 33 исследования, выражающих паттерны сильной презиготической изоляции при симпатрии. Обследование скорости видообразования у рыб и связанных с ними гибридных зон обнаружило сходные закономерности в симпатрии, подтверждающие наличие подкрепления.
Лабораторные исследования, которые явно проверяют наличие подкрепления, ограничены, причем многие эксперименты проводились на плодовых мушках Drosophila. В общем, было проведено два типа экспериментов: с использованием искусственного отбора для имитации естественного отбора, исключающего гибриды (часто называемого «уничтожением гибридов»), и с использованием подрывного отбора для выбора признака ( независимо от его функции в половом размножении). Многие эксперименты с использованием техники уничтожения гибридов обычно упоминаются как поддерживающие подкрепление; однако некоторые исследователи, такие как Койн и Орр, Уильям Р. Райс и Эллен Э. Хостерт, утверждают, что они на самом деле не моделируют подкрепление, поскольку поток генов полностью ограничен между двумя популяциями.
Были предложены различные альтернативные объяснения закономерностей, наблюдаемых в природе. Не существует единого всеобъемлющего признака подкрепления; однако есть две предлагаемые возможности: асимметрия пола (когда женщины в симпатрических популяциях вынуждены становиться разборчивыми перед двумя разными мужчинами) и вариант аллельного доминирования : любой из аллелей, испытывающих отбор на изоляция должна преобладать. Хотя эта сигнатура не полностью учитывает вероятности фиксации или смещение экологического символа. Койн и Орр расширяют признак половой асимметрии и утверждают, что, независимо от изменений, наблюдаемых у женщин и мужчин в симпатии, изоляция в большей степени определяется самками.
Экология также могут играют роль в наблюдаемых закономерностях - это называется смещением экологического характера. Естественный отбор может способствовать сокращению перекрытия ниш между видами вместо того, чтобы уменьшать гибридизацию. Хотя один эксперимент с колюшей рыбой, который явно проверял эту гипотезу, не нашел доказательств.
Взаимодействие видов также может привести к смещению репродуктивного характера (как по предпочтению партнера, так и по сигналу о спаривании). Примеры включают давление хищников и конкуренции, паразиты, обманчивое опыление и мимикрия. Потому что эти и другие факторы могут привести к смещению репродуктивного характера и дают пять критериев для подкрепления, которые следует различать между экологическими и этологическими влияниями:
(1) признаки спаривания идентифицируются у основных видов; (2) на брачные признаки влияет взаимодействие видов, так что отбор по брачным признакам вероятен; (3) виды взаимодействия различаются между популяциями (присутствующие и отсутствующие, или различные виды взаимодействия, влияющие на брачные черты в каждой популяции); (4) брачные признаки (сигнал и / или предпочтение) различаются между популяциями из-за различий во взаимодействии видов; (5) видообразование требует демонстрации того, что дивергенция брачных признаков приводит к полной или почти полной сексуальной изоляции среди популяций. Результаты будут наиболее информативными в хорошо разрешенной биогеографической обстановке, где известны взаимоотношения и история популяций.
Возможно, что модель усиленной изоляции может быть просто временным результатом вторичного контакта где два аллопатрических вида уже имеют различный диапазон презиготической изоляции: одни проявляют больше, чем другие. Те, у кого более слабая презиготическая изоляция, в конечном итоге сливаются, теряя свою отличительность. Эта гипотеза не объясняет того факта, что отдельные виды в аллопатрии, испытывающие постоянный поток генов, не будут различаться по уровням потока генов при вторичном контакте. Более того, паттерны, обнаруженные у Drosophila, обнаруживают высокие уровни презиготической изоляции в симпатрии, но не в аллопатрии. Гипотеза слияния предсказывает, что сильная изоляция должна быть найдена как в аллопатрии, так и в симпатрии. Считается, что этот процесс слияния происходит в природе, но не полностью объясняет закономерности, обнаруживаемые при подкреплении.
 Филогенетическая подпись, позволяющая отличить симпатрическое видообразование от подкрепления. Более сильная презиготическая изоляция (обозначенная красными прямоугольниками и соответствующими стрелками) должна быть обнаружена между Z и Y и между Z и X, если вид Z симпатрически виден (зеленый) от общего предка видов Y и X. Если Z, Y и X видообразование аллопатрически (синий), с Z и Y, испытывающими вторичный контакт, сильная презиготическая изоляция должна быть найдена между Z и Y, но не между Z и X.
Филогенетическая подпись, позволяющая отличить симпатрическое видообразование от подкрепления. Более сильная презиготическая изоляция (обозначенная красными прямоугольниками и соответствующими стрелками) должна быть обнаружена между Z и Y и между Z и X, если вид Z симпатрически виден (зеленый) от общего предка видов Y и X. Если Z, Y и X видообразование аллопатрически (синий), с Z и Y, испытывающими вторичный контакт, сильная презиготическая изоляция должна быть найдена между Z и Y, но не между Z и X. Возможно, что процесс симпатрического видообразования сам по себе может привести к наблюдаемым образцам армирования. Один из методов различения этих двух видов - построить филогенетическую историю вида, поскольку сила презиготической изоляции между группой родственных видов должна различаться в зависимости от того, как они видоизменялись в прошлом. Два других способа определить, происходит ли подкрепление (в отличие от симпатрического видообразования):
В процесс беглости (мало чем отличается от убегающий отбор по рыбам ), отбор против гибридов с низкой приспособленностью способствует ассортивному спариванию, быстро увеличивая дискриминацию спаривания. Кроме того, когда предпочтения женского партнера обходятся недорого, в результате могут происходить изменения мужских фенотипов, выражающие паттерн, идентичный паттерну репродуктивного смещения признаков. Постзиготическая изоляция не нужна, поскольку она вызвана просто тем фактом, что непригодные гибриды не могут получить партнеров.
Был выдвинут ряд возражений, в основном в течение 1980-х, утверждая, что усиление неправдоподобно. Большинство полагается на теоретические работы, которые предполагали, что антагонизм между силами естественного отбора и потоком генов был самым большим препятствием на пути к его выполнимости. Эти возражения с тех пор в значительной степени опровергаются свидетельствами природы.
Обеспокоенность по поводу того, что приспособленность гибридов играет роль в подкреплении, привела к возражениям, основанным на взаимосвязи между отбором и рекомбинацией. То есть, если поток генов не равен нулю (если гибриды не полностью непригодны), отбор не может управлять фиксацией аллелей для презиготической изоляции. Например: если популяция 





Кроме того, определенные аллели, которые имеют селективное преимущество в перекрывающихся популяциях, полезны только внутри этой популяции. Однако, если они имеют селективное преимущество, поток генов должен позволить аллелям распространяться по обеим популяциям. Чтобы предотвратить это, аллели должны быть вредными или нейтральными. Это не без проблем, поскольку поток генов из предположительно больших аллопатрических областей может подавить область, когда две популяции перекрываются. Чтобы подкрепление работало, поток генов должен присутствовать, но очень ограниченный.
Недавние исследования показывают, что подкрепление может происходить в более широком диапазоне условий, чем считалось ранее, и что эффект потока генов можно преодолеть путем отбора. Например, два вида Drosophila santomea и D. якуба на африканском острове Сан-Томе время от времени скрещиваются друг с другом, что приводит к плодовитым потомкам женского пола и бесплодным потомкам мужского пола. Эта естественная обстановка была воспроизведена в лаборатории, непосредственно моделируя подкрепление: удаление некоторых гибридов и допуск различных уровней потока генов. Результаты эксперимента убедительно свидетельствуют о том, что подкрепление работает в различных условиях, с эволюцией половой изоляции, возникающей в 5–10 поколениях плодовых мух.
В сочетании со слиянием Согласно гипотезе подкрепление можно рассматривать как гонку против слияния и вымирания. Получение непригодных гибридов фактически то же самое, что и недостаток гетерозиготы; при этом отклонение от генетического равновесия вызывает потерю непригодного аллеля. Этот эффект приведет к исчезновению одной из популяций. Это возражение преодолевается, когда обе популяции не находятся в одинаковых экологических условиях. Тем не менее, вымирание одной популяции все еще возможно, и это было показано в популяционном моделировании. Чтобы подкрепление произошло, презиготическая изоляция должна произойти быстро.