
Войти
| Часть серии о |
| Эволюционная биология |
|---|
 Зябликов Дарвина по Джон Гулд Зябликов Дарвина по Джон Гулд |
| Процессы и результаты |
| Естественная история |
| История эволюционной теории |
| Поля и приложения |
| Социальные последствия |
|
|
Коэволюция паразита и хозяина - это особый случай совместной эволюции, когда хозяин и паразит постоянно адаптируются друг к другу. Это может создать между ними эволюционную гонку вооружений. Более благоприятная возможность - эволюционный компромисс между передачей и вирулентностью паразита, так как если паразит убивает своего хозяина слишком быстро, паразит также не может воспроизводиться. Другая теория, гипотеза Красной Королевы, предполагает, что, поскольку и хозяин, и паразит должны продолжать развиваться, чтобы не отставать друг от друга, и поскольку половое размножение постоянно создает новые комбинации генов, паразитизм способствует половому размножению в хозяине.
Связанные с этим генетические изменения - это изменения частот аллелей, вариантных форм отдельных генов в популяциях. Они определяются тремя основными типами динамики отбора : отрицательный частотно-зависимый отбор, когда редкий аллель имеет селективное преимущество; преимущество гетерозиготы ; и направленный отбор вблизи выгодного аллеля. Возможный результат - географическая мозаика в паразитированной популяции, поскольку и хозяин, и паразит приспосабливаются к условиям окружающей среды, которые меняются в пространстве и времени.
Совместная эволюция паразита и хозяина широко распространена как в дикой природе, так и среди людей, домашних животных и сельскохозяйственных культур. Основные заболевания, такие как малярия, СПИД и грипп, вызываются одновременным развитием паразитов.
Модельные системы для изучения коэволюции паразита и хозяина включают нематоду Caenorhabditis elegans с бактерией Bacillus thuringiensis ; рачок дафния и его многочисленные паразиты; и бактерия Escherichia coli и млекопитающие (включая человека), в кишечнике которых она обитает.
Хозяева и паразиты оказывают взаимное избирательное давление друг на друга, что может привести к быстрой взаимной адаптации. Для организмов с коротким временем генерации совместную эволюцию паразита и хозяина можно наблюдать в сравнительно короткие периоды времени, что позволяет изучать эволюционные изменения в режиме реального времени как в полевых, так и в лабораторных условиях. Таким образом, эти взаимодействия могут служить противоположным примером общепринятому представлению о том, что эволюцию можно обнаружить только в течение длительного времени.
Динамика этих взаимодействий резюмируется в гипотезе Красной Королевы, а именно, что и хозяин, и паразит должны постоянно меняться, чтобы идти в ногу с адаптациями друг друга.
Совместная эволюция паразита и хозяина является повсеместной и имеет потенциальное значение для всех живых организмов, включая людей, домашних животных и сельскохозяйственных культур. Основные заболевания, такие как малярия, СПИД и грипп, вызываются одновременным развитием паразитов. Лучшее понимание коэволюционной адаптации между стратегиями атаки паразитов и иммунной системой хозяина может помочь в разработке новых лекарств и вакцин.
Коэволюция паразит-хозяин характеризуется реципрокным генетическим изменением и, следовательно, изменениями частот аллелей в популяциях. Эти изменения можно определить по трем основным типам динамики отбора.
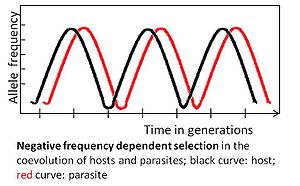
Аллель подвергается отрицательному частотно-зависимому отбору, если редкий аллельный вариант имеет селективное преимущество. Например, паразит должен адаптироваться к наиболее распространенному генотипу хозяина, потому что тогда он может заразить большое количество хозяев. В свою очередь, редкий генотип хозяина может быть одобрен отбором, его частота увеличится и, в конечном итоге, он станет обычным. Впоследствии паразиту следует адаптироваться к бывшему нечастому генотипу.
Коэволюция, определяемая отрицательным частотно-зависимым отбором, происходит быстро и может происходить через несколько поколений. Он поддерживает высокое генетическое разнообразие, отдавая предпочтение необычным аллелям. Этот режим отбора ожидается для многоклеточных хозяев, поскольку адаптации могут происходить без необходимости новых полезных мутаций, которые с меньшей вероятностью будут встречаться у этих хозяев из-за относительно небольшого размера популяции и относительно длительного времени генерации.
Избыточное доминирование возникает, если фенотип гетерозиготы имеет преимущество приспособляемости по сравнению с обоими гомозиготами ( преимущество гетерозиготы, вызывающее гетерозис ). Одним из примеров является серповидноклеточная анемия. Это происходит из-за мутации в гене гемоглобина, приводящей к серповидному образованию красных кровяных телец, вызывая тромбообразование в кровеносных сосудах, ограничение кровотока и снижение транспорта кислорода. В то же время мутация придает устойчивость к малярии, вызываемой паразитами Plasmodium, которые передаются в эритроцитах после передачи комарам человеку. Следовательно, гомозиготные и гетерозиготные генотипы по аллелю серповидно-клеточной анемии проявляют устойчивость к малярии, в то время как гомозигота страдает фенотипом тяжелого заболевания. Альтернативная гомозигота, не несущая аллеля серповидноклеточной анемии, восприимчива к инфицированию Plasmodium. Как следствие, гетерозиготный генотип избирательно предпочитается в районах с высокой заболеваемостью малярией.
 Дополнительная информация: выбор направления и выборочная развертка
Дополнительная информация: выбор направления и выборочная развертка Если аллель дает преимущество в пригодности, его частота увеличивается в популяции - отбор бывает направленным или положительным. Селективный развертки является формой направленного отбора, где увеличение частоты в итоге приводит к фиксации выгодного аллель и других аллелей вблизи него. Считается, что этот процесс идет медленнее, чем отрицательный частотно-зависимый выбор. Это может вызвать «гонку вооружений», состоящую из повторяющегося возникновения и закрепления новых черт вирулентности у паразита и новых защитных черт у хозяина.
Этот способ отбора, вероятно, будет происходить во взаимодействиях между одноклеточными организмами и вирусами из-за больших размеров популяции, короткого времени генерации, часто гаплоидных геномов и горизонтального переноса генов, что увеличивает вероятность возникновения и распространения полезных мутаций в популяциях.
Географическая мозаичная теория коэволюции Джона Н. Томпсона выдвигает гипотезу о пространственно расходящемся коэволюционном отборе, приводящем к генетической дифференциации между популяциями. Модель предполагает три элемента, которые вместе подпитывают коэволюцию:
 Подорожник lanceolata взаимодействует с мучнистой росой (не показана), создавая географическую мозаику генотипов хозяина и паразита.
Подорожник lanceolata взаимодействует с мучнистой росой (не показана), создавая географическую мозаику генотипов хозяина и паразита. 1) мозаика отбора среди популяций
2) коэволюционные горячие точки
3) географическое смешение признаков
Эти процессы интенсивно изучались на растении Plantago lanceolata и его паразите - мучнистой росе Podosphaera plantaginis на Аландских островах в юго-западной Финляндии. Плесень получает питательные вещества от своего хозяина, многолетнего растения, посылая корни в растение. В этом регионе насчитывается более 3000 популяций хозяев, где обе популяции могут свободно развиваться в неоднородном ландшафте в отсутствие отбора, навязанного человеком. Оба партнера могут размножаться бесполым и половым путем. Плесень обычно вымирает на местном уровне зимой, а летом вызывает местные эпидемии. Успешность перезимовки плесени и интенсивность встреч между хозяином и патогеном летом сильно различаются географически. Система имеет пространственно расходящуюся коэволюционную динамику в двух метапопуляциях, как предсказывает теория мозаики.
 Potamopyrgus antipodarum имеет тенденцию к размножению половым путем в присутствиипаразитов- трематод.
Potamopyrgus antipodarum имеет тенденцию к размножению половым путем в присутствиипаразитов- трематод. Гипотеза Красной Королевы утверждает, что и хозяин, и паразит должны постоянно меняться, чтобы идти в ногу с адаптациями друг друга, как это описано в художественной литературе Льюиса Кэрролла. Биолог-математик-эволюционист У. Д. Гамильтон далее предположил, что, поскольку половое размножение постоянно создает новые комбинации генов, некоторые из которых могут быть полезными, паразитизм способствует половому размножению в организме хозяина.
Новозеландская пресноводная улитка Potamopyrgus antipodarum и различные ее паразиты- трематоды представляют собой довольно особую модельную систему. Популяции P. antipodarum состоят из бесполых клонов и половых особей и поэтому могут использоваться для изучения эволюции и преимуществ полового размножения. Существует высокая корреляция между присутствием паразитов и частотой половых людей в различных популяциях. Этот результат согласуется с одобрением полового размножения, предложенным в гипотезе Красной Королевы.
 Tribolium castaneum
Tribolium castaneum Красный мучной жук Tribolium castaneum является хозяином микроспоридий Nosema whitei. Этот паразитоид убивает своего хозяина для передачи, поэтому продолжительность жизни хозяина важна для успеха паразита. В свою очередь, приспособленность паразитов, скорее всего, зависит от компромисса между передачей (споровая нагрузка) и вирулентностью. Более высокая вирулентность увеличит потенциал для производства большего количества потомства, но более высокая нагрузка спор повлияет на продолжительность жизни хозяина и, следовательно, на скорость передачи. Этот компромисс подтверждается коэволюционными экспериментами, которые выявили снижение вирулентности, постоянный потенциал передачи и увеличение продолжительности жизни хозяина в течение определенного периода времени. Дальнейшие эксперименты продемонстрировали более высокую скорость рекомбинации у хозяина во время коэволюционных взаимодействий, что может быть избирательно выгодным, поскольку должно увеличивать разнообразие генотипов хозяина.
Ресурсы обычно ограничены. Таким образом, предпочтение одного признака, такого как вирулентность или иммунитет, ограничивает другие жизненные признаки, такие как репродуктивная способность. Более того, гены часто являются плейотропными и имеют множество эффектов. Таким образом, изменение гена плейотропного иммунитета или вирулентности может автоматически влиять на другие признаки. Таким образом, существует компромисс между преимуществами и затратами адаптивных изменений, которые могут помешать популяции хозяина стать полностью резистентной или популяцией паразита быть высокопатогенными. Стоимость генной плейотропии была исследована при совместной эволюции Escherichia coli и бактериофагов. Чтобы ввести свой генетический материал, фагам необходимо связываться со специфическим рецептором бактериальной клеточной поверхности. Бактерия может предотвратить инъекцию, изменяя соответствующий сайт связывания, например, в ответ на точечные мутации или делецию рецептора. Однако эти рецепторы выполняют важные функции в метаболизме бактерий. Их потеря, таким образом, снизит приспособленность (то есть скорость роста населения). Как следствие, существует компромисс между преимуществами и недостатками мутированного рецептора, что приводит к полиморфизму в этом локусе.
 Caenorhabditis elegans
Caenorhabditis elegans Нематода Caenorhabditis elegans и бактерия Bacillus thuringiensis стали модельной системой для изучения коэволюции паразита и хозяина. Лабораторные эволюционные эксперименты предоставили доказательства многих основных предсказаний этих коэволюционных взаимодействий, включая взаимные генетические изменения, увеличение скорости эволюции и увеличение генетического разнообразия.
 Дафния великая
Дафния великая Ракообразные дафнии и их многочисленные паразиты стали одной из основных модельных систем для изучения коэволюции. Хозяин может быть как бесполым, так и половым (вызванным изменениями во внешней среде), поэтому половое размножение можно стимулировать в лаборатории. Десятилетия совместной эволюции между Daphnia magna и бактерией Pasteuria ramosa были реконструированы, реанимируя стадии покоя обоих видов из слоистых отложений пруда и подвергая хозяев из каждого слоя паразитам из прошлого, того же самого и будущих слоев. Исследование показало, что паразиты в среднем наиболее заразны со своими современными хозяевами, что соответствует отрицательному частотно-зависимому отбору.
 кишечная палочка
кишечная палочка Escherichia coli, грамотрицательная протеобактерия, является распространенной моделью в биологических исследованиях, для которых доступны исчерпывающие данные о различных аспектах ее жизненного цикла. Он широко используется для эволюционных экспериментов,том числе связанного с коэволюция с фагами. Эти исследования показали, среди прочего, что на коэволюционную адаптацию могут влиять плейотропные эффекты вовлеченных генов. В частности, связывание бактериофага споверхностным рецептором E. coli является решающим этапом вцикле вирусной инфекции. Мутации в сайте связывания рецептора может вызвать сопротивление. Такие мутации часто проявляют плейотропные эффекты и могут вызвать сопротивление. В присутствии фагов такая плейотропия может приводить к полиморфизму в бактериальной популяции и, таким образом, увеличивать биоразнообразие в сообществе.
Другая модельная система состоит из бактерии Pseudomonas, колонизирующей растения и животных, и ее бактериофагов. Эта система позволила по-новому взглянуть на динамику коэволюционных изменений. Он продемонстрировал, что коэволюция может происходить через повторяющиеся выборочные зачистки, в пользу генералистов для обоих партнеров. Кроме того, совместная эволюция с фагами может способствовать аллопатрическому разнообразию, потенциально увеличивая биоразнообразие и, возможно, видообразование. Совместная эволюция паразита и хозяина также может влиять на лежащую в основе генетику, например, способствуя увеличению частоты мутаций у хозяина.
Взаимодействие тропических деревьев и лиан также было предметом изучения. Здесь лианы рассматривались как сверхразнообразные универсальные макропаразиты, которые влияют на выживание хозяина, паразитируя на его структурной поддержке для доступа к свету навеса, узурпируя ресурсы, которые в противном случае были бы доступны их хозяину. Деревья-хозяева имеют самые разные уровни устойчивости к заражению крон лианами.