
Войти
 Типичная эукариотическая клетка
Типичная эукариотическая клетка Клеточное дыхание - это набор метаболических реакций и процессов, которые происходят в клетках организмов для преобразования химической энергии из молекул кислорода или питательных веществ в аденозинтрифосфат (АТФ) с последующим выделением отходов. Реакции, участвующие в дыхании, - это катаболические реакции, которые разбивают большие молекулы на более мелкие, высвобождая энергию, потому что слабые высокоэнергетические связи, в частности в молекулярном кислороде, заменяются более прочными связями в продуктах. Дыхание - один из ключевых способов высвобождения химической энергии клеткой для подпитки клеточной активности. Общая реакция происходит в виде серии биохимических стадий, некоторые из которых являются окислительно-восстановительными реакциями. Хотя клеточное дыхание технически является реакцией горения, оно явно не похоже на то, что происходит в живой клетке, из-за медленного контролируемого высвобождения энергии из серии реакций.
Питательные вещества, которые обычно используются клетками животных и растений при дыхании, включают сахар, аминокислоты и жирные кислоты, а наиболее распространенные окислитель, обеспечивающий большую часть химической энергии, представляет собой молекулярный кислород (O2). Химическая энергия, запасенная в АТФ (связь его третьей фосфатной группы с остальной частью молекулы может быть разорвана, позволяя формировать более стабильные продукты, тем самым высвобождая энергию для использования клеткой), затем может использоваться для управления процессами, требующими энергии, включая биосинтез, перемещение или перенос молекул через клеточные мембраны.
Аэробное дыхание требует кислорода (O2) для создания АТФ. Хотя углеводы, жиры и белки потребляются в качестве реагентов, аэробное дыхание является предпочтительным методом разложения пирувата при гликолизе., и требует пирувата для митохондрий, чтобы полностью окислиться с помощью цикла лимонной кислоты. Продуктами этого процесса являются диоксид углерода и вода, а передаваемая энергия используется для разрыва связей в АДФ с целью добавления третьей фосфатной группы с образованием АТФ (аденозинтрифосфат ) посредством фосфорилирования на уровне субстрата., NADH и FADH 2
| Упрощенная реакция: | C6H12O6(s) + 6 O 2 (g) → 6 CO 2 (г) + 6 H 2 O (л) + тепло |
| ΔG = -2880 кДж на моль C 6H12O6 |
Отрицательный ΔG указывает, что реакция может происходить самопроизвольно.
Потенциал НАДН и ФАДН 2 преобразуется в большее количество АТФ через цепь переноса электронов с кислородом и протонами (водородом) в качестве «концевых акцепторов электронов». Большая часть АТФ, продуцируемого аэробным клеточным дыханием, образуется за счет окислительного фосфорилирования. Выделяемая энергия O 2 используется для создания хемиосмотического потенциала путем прокачки протонов через мембрану. Этот потенциал затем используется для управления АТФ-синтазой и производства АТФ из АДФ и фосфатной группы. В учебниках биологии часто говорится, что 38 молекул АТФ могут образовываться на одну окисленную молекулу глюкозы во время клеточного дыхания (2 из гликолиза, 2 из цикла Кребса и около 34 из системы транспорта электронов). Однако этот максимальный выход никогда не достигается полностью из-за потерь из-за протекающих мембран, а также из-за затрат на перемещение пирувата и АДФ в митохондриальный матрикс, и текущие оценки колеблются в пределах от 29 до 30 АТФ на глюкозу.
Аэробные нагрузки. метаболизм до 15 раз более эффективен, чем анаэробный метаболизм (который дает 2 молекулы АТФ на 1 молекулу глюкозы), потому что двойная связь в O 2 имеет более высокую энергию, чем другие двойные связи или пары одинарных связей в других общие молекулы в биосфере. Однако некоторые анаэробные организмы, такие как метаногены, могут продолжать анаэробное дыхание, производя больше АТФ за счет использования других неорганических молекул (не кислорода) с высокой энергией в качестве конечных акцепторов электронов в электронная транспортная цепь. Они разделяют начальный путь гликолиза, но аэробный метаболизм продолжается с циклом Кребса и окислительным фосфорилированием. Постгликолитические реакции происходят в митохондриях эукариотических клеток и в цитоплазме в прокариотических клетках.
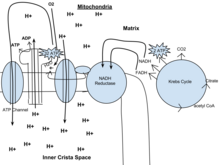 Из цитоплазмы он переходит в цикл Кребса с ацетил-КоА. Затем он смешивается с CO 2 и образует 2 АТФ, NADH и FADH. Оттуда НАДН и ФАДН переходят в НАДН-редуктазу, которая производит фермент. НАДН тянет электроны фермента, чтобы послать их через цепь переноса электронов. Цепь переноса электронов тянет ионы H через цепь. Из цепи переноса электронов высвободившиеся ионы водорода производят АДФ с конечным результатом 32 АТФ. O 2 обеспечивает большую часть энергии для процесса и объединяется с протонами и электронами, образуя воду. Наконец, АТФ выходит через АТФ-канал и выходит из митохондрий.
Из цитоплазмы он переходит в цикл Кребса с ацетил-КоА. Затем он смешивается с CO 2 и образует 2 АТФ, NADH и FADH. Оттуда НАДН и ФАДН переходят в НАДН-редуктазу, которая производит фермент. НАДН тянет электроны фермента, чтобы послать их через цепь переноса электронов. Цепь переноса электронов тянет ионы H через цепь. Из цепи переноса электронов высвободившиеся ионы водорода производят АДФ с конечным результатом 32 АТФ. O 2 обеспечивает большую часть энергии для процесса и объединяется с протонами и электронами, образуя воду. Наконец, АТФ выходит через АТФ-канал и выходит из митохондрий. Гликолиз - это метаболический путь, который происходит в цитозоле клеток всех живых организмов. Гликолиз можно буквально перевести как «расщепление сахара», он происходит в присутствии кислорода или без него. В аэробных условиях процесс превращает одну молекулу глюкозы в две молекулы пирувата (пировиноградной кислоты), генерируя энергию в виде двух чистых молекул АТФ. Фактически производятся четыре молекулы АТФ на глюкозу, однако две из них потребляются как часть подготовительной фазы. Первоначальное фосфорилирование глюкозы требуется для увеличения реакционной способности (снижения ее стабильности), чтобы молекула была расщеплена на две молекулы пирувата ферментом альдолазой. Во время фазы отдачи гликолиза четыре фосфатные группы переносятся на АДФ посредством фосфорилирования на уровне субстрата с образованием четырех АТФ, и два НАДН образуются, когда пируват окисляется. Общая реакция может быть выражена следующим образом:
Начиная с глюкозы, 1 АТФ используется для передачи фосфата глюкозе с образованием глюкозо-6-фосфата. Гликоген может быть преобразован в глюкозо-6-фосфат также с помощью гликогенфосфорилазы. Во время энергетического обмена глюкозо-6-фосфат становится фруктозо-6-фосфатом. Дополнительный АТФ используется для фосфорилирования фруктозо-6-фосфата в фруктозо-1,6-бисфосфат с помощью фосфофруктокиназы. Затем 1,6-бифосфат фруктозы расщепляется на две фосфорилированные молекулы с тремя углеродными цепями, которые позже распадаются на пируват.
Пируват окисляется до ацетил-КоА и CO 2 с помощью пируватдегидрогеназного комплекса (PDC). PDC содержит несколько копий трех ферментов и расположен в митохондриях эукариотических клеток и в цитозоле прокариот. При превращении пирувата в ацетил-КоА образуется одна молекула НАДН и одна молекула СО 2.
Это также называется циклом Кребса или циклом трикарбоновой кислоты. Когда присутствует кислород, ацетил-КоА образуется из молекул пирувата, образованных в результате гликолиза. После образования ацетил-КоА может происходить аэробное или анаэробное дыхание. Когда присутствует кислород, митохондрии подвергаются аэробному дыханию, которое приводит к циклу Кребса. Однако, если кислорода нет, происходит ферментация молекулы пирувата. В присутствии кислорода, когда образуется ацетил-КоА, молекула затем входит в цикл лимонной кислоты (цикл Кребса) внутри митохондриального матрикса и окисляется до CO2, в то же время восстанавливая NAD до NADH. НАДН может использоваться цепью переноса электронов для создания дополнительного АТФ как части окислительного фосфорилирования. Чтобы полностью окислить эквивалент одной молекулы глюкозы, два ацетил-КоА должны метаболизироваться в цикле Кребса. В течение этого цикла образуются два низкоэнергетических отходов, H 2 O и CO 2.
Цикл лимонной кислоты - это 8-этапный процесс, в котором участвуют 18 различных ферментов и коферментов. Во время цикла ацетил-КоА (2 атома углерода) + оксалоацетат (4 атома углерода) дает цитрат (6 атомов углерода), который превращается в более реактивную форму, называемую изоцитратом (6 атомов углерода). Изоцитрат модифицируется в α-кетоглутарат (5 атомов углерода), сукцинил-КоА, сукцинат, фумарат, малат и, наконец, оксалоацетат.
Чистый выигрыш от одного цикла составляет 3 НАДН и 1 ФАДН 2 в качестве соединений, переносящих водород (протон плюс электрон), и 1 высокоэнергетический GTP, который впоследствии может быть использован для получения АТФ. Таким образом, общий выход из 1 молекулы глюкозы (2 молекул пирувата) составляет 6 НАДН, 2 ФАДН 2 и 2 АТФ.
У эукариот окислительное фосфорилирование происходит в митохондриальных кристах. Он включает цепь переноса электронов, которая устанавливает протонный градиент (хемиосмотический потенциал) через границу внутренней мембраны за счет окисления НАДН, образующегося в цикле Кребса. АТФ синтезируется ферментом АТФ-синтаза, когда хемиосмотический градиент используется для управления фосфорилированием АДФ. Перенос электронов осуществляется за счет химической энергии экзогенного кислорода, и при добавлении двух протонов образуется вода.
В таблице ниже описаны реакции, происходящие, когда одна молекула глюкозы полностью окисляется до диоксида углерода. Предполагается, что все восстановленные коферменты окисляются цепью переноса электронов и используются для окислительного фосфорилирования.
| Этап | выход кофермента | Выход АТФ | Источник АТФ |
|---|---|---|---|
| Подготовительная фаза гликолиза | -2 | Фосфорилирование глюкозы и фруктозо-6-фосфат использует два АТФ из цитоплазмы. | |
| Фаза возврата гликолиза | 4 | Фосфорилирование на уровне субстрата | |
| 2 NADH | 3 или 5 | Окислительное фосфорилирование: каждый NADH производит чистое 1,5 АТФ (вместо обычных 2,5) за счет транспорта НАДН через митохондриальную мембрану | |
| Окислительное декарбоксилирование пирувата | 2 НАДН | 5 | Окислительное фосфорилирование |
| Цикл Кребса | 2 | Фосфорилирование на уровне субстрата | |
| 6 НАДН | 15 | Окислительное фосфорилирование | |
| 2 FADH 2 | 3 | Окислительное фосфорилирование | |
| Общий выход | 30 или 32 АТФ | От полного окисления одной молекулы глюкозы до диоксида углерода и окисления всего восстановленные коферменты. | |
Хотя существует теоретический выход 38 молекул АТФ на глюкозу во время клеточного дыхания, такие условия обычно не достигаются из-за потерь, таких как стоимость перемещения пирувата (из гликолиза), фосфата и АДФ (субстратов для синтеза АТФ) в митохондрии. Все они активно транспортируются с помощью носителей, которые используют накопленную энергию в протонном электрохимическом градиенте..
Результат этих транспортных процессов с использованием протонный электрохимический градиент состоит в том, что для образования 1 АТФ необходимо более 3 H. Очевидно, это снижает теоретическую эффективность всего процесса, и вероятный максимум приближается к 28–30 молекулам АТФ. На практике эффективность может быть даже ниже, потому что внутренняя мембрана митохондрий слегка пропускает протоны. Другие факторы также могут рассеивать протонный градиент, создавая явно протекающие митохондрии. Несвязывающий белок, известный как термогенин, экспрессируется в некоторых типах клеток и является каналом, по которому могут транспортироваться протоны. Когда этот белок активен во внутренней мембране, он замыкает связь между цепью переноса электронов и синтезом АТФ. Потенциальная энергия протонного градиента не используется для производства АТФ, но генерирует тепло. Это особенно важно для термогенеза бурого жира новорожденных и гибернирующих млекопитающих.
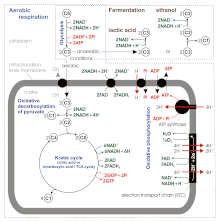 Стехиометрия аэробного дыхания и наиболее известных типов ферментации в эукариотической клетке. Цифры в кружках указывают количество атомов углерода в молекулах, C6 - это глюкоза C6H12O6, C1 углекислый газ CO2. митохондриальная внешняя мембрана не указана.
Стехиометрия аэробного дыхания и наиболее известных типов ферментации в эукариотической клетке. Цифры в кружках указывают количество атомов углерода в молекулах, C6 - это глюкоза C6H12O6, C1 углекислый газ CO2. митохондриальная внешняя мембрана не указана. Согласно некоторым новым источникам, АТФ выход во время аэробного дыхания составляет не 36–38, а всего лишь около 30–32 молекул АТФ / 1 молекулу глюкозы, потому что:
Итак, наконец, мы имеем на молекулу глюкозы
В сумме это дает 4 + 3 (или 5) + 20 + 3 = 30 (или 32) АТФ на молекулу глюкозы
Эти цифры могут потребовать дополнительной настройки по мере появления новых структурных деталей. Приведенное выше значение 3 H + / ATP для синтазы предполагает, что синтаза перемещает 9 протонов и производит 3 ATP за один оборот. Количество протонов зависит от количества с-субъединиц в с-кольце Fo, и теперь известно, что оно составляет 10 у дрожжей Fo и 8 у позвоночных. Включая один H + для транспортных реакций, это означает, что для синтеза одного АТФ требуется 1 + 10/3 = 4,33 протона у дрожжей и 1 + 8/3 = 3,67 у позвоночных. Это означало бы, что в митохондриях человека 10 протонов от окисления НАДН будут производить 2,72 АТФ (вместо 2,5), а 6 протонов от окисления сукцината или убихинола будут производить 1,64 АТФ (вместо 1,5). Это согласуется с экспериментальными результатами в пределах погрешности, описанными в недавнем обзоре.
Общий выход АТФ в этаноле или молочной кислоте ферментация составляет всего 2 молекулы, полученные в результате гликолиза, поскольку пируват не переносится в митохондрию и, наконец, не окисляется до диоксида углерода (CO 2), а восстанавливается до этанола или молочной кислоты. кислота в цитоплазме.
Без кислорода пируват (пировиноградная кислота ) не метаболизируется посредством клеточного дыхания, но подвергается процессу ферментации. Пируват не транспортируется в митохондрии, но остается в цитоплазме, где он превращается в продукты жизнедеятельности, которые можно удалить из клетки. Это служит цели окисления переносчиков электронов, чтобы они снова могли выполнять гликолиз и удалять избыток пирувата. Ферментация окисляет НАДН до НАД, поэтому его можно повторно использовать в гликолизе. В отсутствие кислорода ферментация предотвращает накопление НАДН в цитоплазме и обеспечивает НАД для гликолиза. Эти отходы различаются в зависимости от организма. В скелетных мышцах продуктом жизнедеятельности является молочная кислота. Этот тип ферментации называется молочно-кислотным брожением. При напряженных упражнениях, когда потребность в энергии превышает запас энергии, дыхательная цепь не может обработать все атомы водорода, к которым присоединен NADH. Во время анаэробного гликолиза НАД регенерируется, когда пары водорода соединяются с пируватом с образованием лактата. Образование лактата катализируется лактатдегидрогеназой в обратимой реакции. Лактат также может использоваться в качестве косвенного предшественника гликогена печени. Во время восстановления, когда становится доступным кислород, НАД присоединяется к водороду из лактата с образованием АТФ. В дрожжах отходами являются этанол и диоксид углерода. Этот тип ферментации известен как спиртовое или этанольное брожение. АТФ, образующийся в этом процессе, производится путем фосфорилирования на уровне субстрата, для которого не требуется кислород.
Ферментация менее эффективна при использовании энергии глюкозы: только 2 АТФ вырабатываются на глюкозу по сравнению с 38 АТФ на глюкозу, номинально производимую при аэробном дыхании. Это связано с тем, что большая часть энергии аэробного дыхания поступает из O 2 с его относительно слабой высокоэнергетической двойной связью. Однако гликолитический АТФ создается быстрее. Чтобы прокариоты продолжали быстро расти, когда они переходят из аэробной среды в анаэробную, они должны увеличивать скорость гликолитических реакций. Для многоклеточных организмов во время коротких всплесков напряженной активности мышечные клетки используют ферментацию, чтобы дополнить выработку АТФ из-за более медленного аэробного дыхания, поэтому ферментация может использоваться клеткой даже до того, как уровень кислорода истощится, как это имеет место в спорте. не требуют от спортсменов скорости, например, спринт.
Клеточное дыхание - это процесс, при котором биологическое топливо окисляется в присутствии высокоэнергетического неорганического акцептора электронов (например, кислорода) для производства большого количества энергии, чтобы стимулировать массовое производство АТФ.
Анаэробное дыхание используется некоторыми микроорганизмами, у которых ни кислород (аэробное дыхание), ни производные пирувата (ферментация) не являются конечными высокоэнергетическими акцепторами электронов. Скорее, используется неорганический акцептор, такой как сульфат или нитрат. Такие организмы обычно встречаются в необычных местах, таких как подводные пещеры или около гидротермальных источников на дне океана.
В июле 2019 года научное исследование Кидд Майн в Канаде обнаружило дышащие серой организмы, которые живут на глубине 7900 футов под поверхностью и которые дышат серой, чтобы выжить. Эти организмы также примечательны тем, что они потребляют в качестве источника пищи такие минералы, как пирит.