
Войти
 Грибные тела, видимые в мозге дрозофилы в виде двух стеблей. Из Jenett et al., 2006.
Грибные тела, видимые в мозге дрозофилы в виде двух стеблей. Из Jenett et al., 2006. В грибных телах или корпуса pedunculata являются парой структур в мозге от насекомых, других членистоногих, а некоторые аннелиды ( в частности, ragworm Platynereis dumerilii ). Также известно, что они играют роль в обонятельном обучении и памяти. У большинства насекомых грибовидные тела и боковой рог являются двумя верхними областями мозга, которые получают обонятельную информацию от антеннальной доли через проекционные нейроны. Впервые они были идентифицированы и описаны французским биологом Феликсом Дюжарденом в 1850 году.
Грибовидные тела обычно описываются как нейропили, т. Е. Как плотные сети нейрональных отростков ( дендритные и аксонные окончания ) и глии. Они получили свое название от их примерно полусферической чашечки, выступа, соединенного с остальной частью мозга центральным нервным трактом или ножкой.
Большинство наших текущих знаний о грибовидных телах основано на исследованиях нескольких видов насекомых, особенно таракана Periplaneta americana, медоносной пчелы Apis mellifera, саранчи и плодовой мухи Drosophila melanogaster. Исследования грибовидных тел плодовых мух были особенно важны для понимания генетической основы функционирования грибовидных тел, поскольку их геном секвенирован и существует огромное количество инструментов для управления экспрессией их генов.
В мозге насекомого цветоножки грибовидных тел проходят через средний мозг. Они в основном состоят из длинных, плотно упакованных нервных волокон клеток Кеньона, внутренних нейронов грибовидных тел. Эти клетки были обнаружены в грибовидных телах всех исследованных видов, хотя их количество варьируется. Например, у плодовых мушек их около 2500, а у тараканов - около 200000.
Грибовидные тела наиболее известны своей ролью в ассоциативном обучении обоняния. Эти обонятельные сигналы поступают от дофаминергических, октопаминергических, холинергических, серотонинергических и ГАМКергических нейронов вне МБ. Они самые большие у перепончатокрылых, которые, как известно, особенно тщательно контролируют обонятельное поведение. Однако, поскольку грибовидные тела также встречаются у аносмических примитивных насекомых, их роль, вероятно, выходит за рамки обонятельной обработки. Анатомические исследования предполагают роль в обработке визуальных и механосенсорных данных у некоторых видов. В частности, у перепончатокрылых субрегионы нейропиля грибовидного тела специализируются на получении обонятельных, зрительных или обоих типов сенсорной информации. У перепончатокрылых обонятельные органы располагаются в чашечках. У муравьев можно различить несколько слоев, соответствующих различным кластерам клубочков в лепестках антенн, возможно, для обработки различных классов запахов. Есть две основные группы проекционных нейронов, разделяющих антеннальную долю на две основные области: переднюю и заднюю. Группы проекционных нейронов разделены, иннервируя группы клубочков по отдельности и отправляя аксоны отдельными маршрутами, либо через медиально-антенно-протоцеребральный тракт (m-APT), либо через латерально-антенно-протоцеребральный тракт (l-APT), и соединяются двумя слоями в чашечки грибовидных тел. В этих слоях топографически представлена организация двух эфферентных областей антеннальной доли, устанавливающая грубую одотопную карту антеннальной доли в области губы грибовидных тел.
Известно, что грибовидные тела участвуют в обучении и памяти, особенно в отношении обоняния, и поэтому в настоящее время являются предметом интенсивных исследований. Исследования показывают, что у более крупных насекомых грибовидные тела обладают и другими функциями обучения и памяти, такими как ассоциативная память, сенсорная фильтрация, моторный контроль и местная память. Исследования показывают, что грибовидные тела обычно действуют как своего рода детектор совпадений, интегрируя мультимодальные входные данные и создавая новые ассоциации, тем самым предполагая их роль в обучении и памяти. Недавняя работа также показывает доказательства участия грибовидного тела во врожденном обонятельном поведении через взаимодействие с боковым рогом, возможно, с использованием частично стереотипных сенсорных реакций выходных нейронов грибовидного тела (MBON) у разных людей. Хотя связи между проекционными нейронами и клетками Kenyon являются случайными (т.е. не стереотипными для отдельных людей), стереотипия в ответах MBON становится возможной благодаря плотной конвергенции многих клеток Kenyon в несколько MBON наряду с другими свойствами сети.
Информация об запахах может быть закодирована в грибовидном теле с помощью характеристик реагирующих нейронов, а также времени их спайков. Эксперименты на саранче показали, что активность клеток Кеньона синхронизирована с нервными колебаниями с частотой 20 Гц, и они особенно чувствительны к спайкам проекционных нейронов в определенных фазах колебательного цикла.
Нейроны, которые получают сигналы от серотонинергических и ГАМКергических нейронов вне MB, вызывают бодрствование, а экспериментальная стимуляция этих серотонинергических нейронов, расположенных выше по течению, вызывает сон. Целевые нейроны в МБ ингибируются серотонином, ГАМК и их комбинацией. С другой стороны, октопамин не влияет на функцию сна МБ.
 Доли грибовидного тела D. melanogaster с видимыми α / β, α '/ β' и γ нейронами. От Дэвиса, 2011 г.
Доли грибовидного тела D. melanogaster с видимыми α / β, α '/ β' и γ нейронами. От Дэвиса, 2011 г. Мы знаем, что структуры грибовидного тела важны для обонятельного обучения и памяти у дрозофилы, потому что их удаление разрушает эту функцию. Грибовидное тело также способно сочетать информацию о внутреннем состоянии тела и обонятельной информации для определения врожденного поведения. Точные роли конкретных нейронов, составляющих грибовидные тела, до сих пор неясны. Однако эти структуры широко изучаются, потому что многое известно об их генетической структуре. Есть три конкретных класса нейронов, которые составляют доли грибовидного тела: α / β, α '/ β' и γ нейроны, каждый из которых имеет различную экспрессию генов. Тема текущего исследования - какие из этих субструктур грибовидного тела участвуют в каждой фазе и процессе обучения и памяти. Грибовидные тела дрозофилы также часто используются для изучения обучения и памяти, и ими манипулируют из-за их относительно дискретной природы. Обычно тесты обонятельного обучения состоят из раздельного воздействия на мух двух запахов; один связан с импульсами электрического удара ( условный раздражитель, или CS +), а второй - нет ( безусловный раздражитель, или УЗИ). После этого периода обучения мух помещают в Т-образный лабиринт с двумя запахами, размещаемыми по отдельности на обоих концах горизонтальных Т-образных ветвей. Рассчитан процент мух, избегающих CS +, при этом высокий уровень избегания считается свидетельством обучения и памяти.
Недавние исследования, сочетающие кондиционирование запаха и визуализацию клеток, выявили шесть следов памяти, которые совпадают с молекулярными изменениями в обонятельной системе дрозофилы. Три из этих следов связаны с ранним формированием поведенческой памяти. Один такой след был визуализирован в антенной доле (AL) репортерными молекулами синапто-pHluorin. Сразу после кондиционирования дополнительный набор проекционных нейронов в наборе из восьми клубочков в AL становится синаптически активированным условным запахом и сохраняется всего 7 минут. Второй след обнаруживается по экспрессии GCaMP и, таким образом, по увеличению притока Ca 2+ в аксоны α '/ β' нейронов грибовидного тела. Это более продолжительный след, который сохраняется до одного часа после кондиционирования. Третий след памяти - снижение активности переднего парного латерального нейрона, который действует как супрессор формирования памяти через один из его ингибирующих ГАМКергических рецепторов. Снижение кальциевого ответа нейронов APL и последующее снижение высвобождения ГАМК на грибовидные тела сохранялось до 5 минут после кондиционирования запаха.
Промежуточный след памяти зависит от экспрессии гена amn, локализованного в дорсальных парных медиальных нейронах. Увеличение притока кальция и синаптического выброса, которое иннервирует грибовидные тела, становится заметным примерно через 30 минут после сочетания электрического шока с запахом и сохраняется в течение как минимум часа. Обе карты долговременной памяти, которые были нанесены на карту, зависят от активности и синтеза белка CREB и CaMKII и существуют только после пространственного кондиционирования. Первый след обнаруживается в α / β нейронах между 9 и 24 часами после кондиционирования и характеризуется увеличением притока кальция в ответ на условный запах. Второй след долговременной памяти формируется в γ-грибовидных телах и обнаруживается по увеличению притока кальция между 18 и 24 часами после кондиционирования.
Циклический аденозинмонофосфат (цАМФ или циклический АМФ) является вторым мессенджером, который участвует в облегчении притока кальция в нейроны грибовидного тела Drosophila melanogaster. Повышение цАМФ вызывает пресинаптическую пластичность у дрозофилы. На уровни цАМФ влияют как нейротрансмиттеры, такие как дофамин и октопамин, так и сами запахи. Дофамин и октопамин выделяются интернейронами грибовидного тела, в то время как запахи напрямую активируют нейроны в обонятельном пути, вызывая приток кальция через потенциалзависимые кальциевые каналы.
В классической парадигме кондиционирования спаренная деполяризация нейронов (через нанесение ацетилхолина для представления запаха или CS ) с последующим применением дофамина (для представления шока или УЗИ ) приводит к синергетическому увеличению цАМФ в долях грибовидного тела. Эти результаты предполагают, что доли грибовидного тела являются критическим местом интеграции CS / US через действие цАМФ. Этот синергетический эффект первоначально наблюдался при аплизии, где сочетание притока кальция с активацией передачи сигналов G-белка серотонином генерирует аналогичное синергетическое увеличение цАМФ.
Кроме того, это синергетическое увеличение цАМФ опосредовано и зависит от аденилатциклазы брюквы (rut AC), которая чувствительна как к кальцию (что является результатом открытия потенциалзависимых кальциевых каналов запахами), так и к стимуляции G-белка (вызываемой дофамином). В то время как прямое сочетание деполяризации нейронов и дофамина (ацетилхолин, за которым следует дофамин) приводит к синергетическому увеличению цАМФ, прямое объединение деполяризации нейронов и октопамина оказывает субаддитивный эффект на цАМФ. В частности, это означает, что эта пара производит значительно меньше цАМФ, чем сумма каждого стимула по отдельности в долях. Следовательно, rut AC в нейронах грибовидного тела работает как детектор совпадений с дофамином и октопамином, действующими двунаправленно, чтобы влиять на уровни цАМФ.
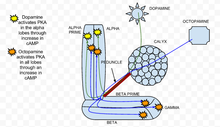 Пространственная регуляция динамики PKA в грибовидном теле дрозофилы.
Пространственная регуляция динамики PKA в грибовидном теле дрозофилы. Было обнаружено, что протеинкиназа A (PKA) играет важную роль в обучении и памяти у дрозофилы. Когда кальций проникает в клетку и связывается с кальмодулином, он стимулирует аденилатциклазу (AC), которая кодируется геном брюквы ( rut). Эта активация AC увеличивает концентрацию цАМФ, который активирует PKA. Когда применяется дофамин, вызывающий отвращение обонятельный стимулятор, он активирует PKA, в частности, в вертикальных долях грибовидного тела. Эта пространственная специфичность регулируется dunce ( dnc) PDE, цАМФ-специфической фосфодиэстеразой. Если ген dunce отменен, как обнаружено у мутанта dnc, пространственная специфичность не сохраняется. Напротив, стимуляция аппетита, создаваемая применением октопамина, увеличивает ПКА во всех долях. В мутанте гона - генотипе, в котором брюква отсутствует, - ответы как на дофамин, так и на октопамин были значительно снижены и близки к экспериментальному шуму.
Ацетилхолин, который представляет собой условный раздражитель, приводит к сильному увеличению активации ПКА по сравнению со стимуляцией только дофамином или октопамином. Эта реакция отсутствует у мутантов рут, что демонстрирует, что PKA важна для сенсорной интеграции. Специфичность активации альфа-доли в присутствии дофамина сохраняется, когда дофамин находится в сочетании с ацетилхолином. По сути, во время парадигмы кондиционирования, когда условный стимул сочетается с безусловным стимулом, PKA проявляет повышенную активацию. Это показывает, что PKA необходима для условного обучения у Drosophila melanogaster.
Стимул → выходные реакции являются продуктом пар возбуждения и торможения. Это тот же образец организации, что и у млекопитающих. Эти паттерны могут, как и у млекопитающих, предшествовать действию. По состоянию на 2021 год это область, только недавно выясненная Zwaka et al 2018, Duer et al 2015 и Paffhausen et al 2020.