
Войти
| индуцируемый гипоксией фактор 1, альфа-субъединица | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Условное обозначение | HIF1A | ||||||
| Ген NCBI | 3091 | ||||||
| HGNC | 4910 | ||||||
| OMIM | 603348 | ||||||
| RefSeq | NM_001530 | ||||||
| UniProt | Q16665 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 14 q21-q24 | ||||||
| |||||||
| Ядерный транслокатор арилуглеводородного рецептора | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Условное обозначение | ARNT | ||||||
| Альт. символы | HIF1B, bHLHe2 | ||||||
| Ген NCBI | 405 | ||||||
| HGNC | 700 | ||||||
| OMIM | 126110 | ||||||
| RefSeq | NM_001668 | ||||||
| UniProt | P27540 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 1 квартал 21 | ||||||
| |||||||
| эндотелиальный белок 1 домена PAS | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Условное обозначение | EPAS1 | ||||||
| Альт. символы | HIF2A, MOP2, PASD2, HLF | ||||||
| Ген NCBI | 2034 г. | ||||||
| HGNC | 3374 | ||||||
| OMIM | 603349 | ||||||
| RefSeq | NM_001430 | ||||||
| UniProt | Q99814 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 2 п.21-п16 | ||||||
| |||||||
| Ядерный транслокатор 2 арил-углеводородного рецептора | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Условное обозначение | ARNT2 | ||||||
| Альт. символы | HIF2B, KIAA0307, bHLHe1 | ||||||
| Ген NCBI | 9915 | ||||||
| HGNC | 16876 | ||||||
| OMIM | 606036 | ||||||
| RefSeq | NM_014862 | ||||||
| UniProt | Q9HBZ2 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 1 квартал 24 | ||||||
| |||||||
| индуцируемый гипоксией фактор 3, альфа-субъединица | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Условное обозначение | HIF3A | ||||||
| Ген NCBI | 64344 | ||||||
| HGNC | 15825 | ||||||
| OMIM | 609976 | ||||||
| RefSeq | NM_152794 | ||||||
| UniProt | Q9Y2N7 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 19 q13 | ||||||
| |||||||
Факторы, индуцируемые гипоксией ( HIF), представляют собой факторы транскрипции, которые реагируют на снижение доступного кислорода в клеточной среде или на гипоксию.
Транскрипционный комплекс HIF был открыт в 1995 году Греггом Л. Семенза и научным сотрудником Гуан Ван. В 2016 году Уильям Кэлин-младший, Питер Дж. Рэтклифф и Грегг Л. Семенца были награждены премией Ласкера за их работу по разъяснению роли HIF-1 в восприятии кислорода и его роли в выживании в условиях низкого содержания кислорода. В 2019 году эти же три человека были совместно удостоены Нобелевской премии по физиологии и медицине за их работу по выяснению того, как HIF воспринимает и адаптирует клеточный ответ к доступности кислорода.
Большинство, если не все виды, дышащие кислородом, экспрессируют высококонсервативный транскрипционный комплекс HIF-1, который представляет собой гетеродимер, состоящий из альфа- и бета-субъединиц, причем последняя является конститутивно экспрессируемым ядерным транслокатором рецептора арильных углеводородов (ARNT). HIF-1 принадлежит к подсемейству PER-ARNT-SIM (PAS) основного семейства факторов транскрипции спираль-петля-спираль (bHLH). Альфа- и бета-субъединица похожи по структуре и обе содержат следующие домены:
|
| ||||||||||||||||||||||||||||||||||||||||||||||
Ниже перечислены члены семьи HIF человека:
| Член | Ген | Протеин |
|---|---|---|
| HIF-1α | HIF1A | индуцируемый гипоксией фактор 1, альфа-субъединица |
| HIF-1β | ARNT | Ядерный транслокатор арилуглеводородного рецептора |
| HIF-2α | EPAS1 | эндотелиальный белок 1 домена PAS |
| HIF-2β | ARNT2 | ядерный транслокатор 2 арил-углеводородного рецептора |
| HIF-3α | HIF3A | фактор, индуцируемый гипоксией 3, альфа-субъединица |
| HIF-3β | ARNT3 | Ядерный транслокатор арил-углеводородного рецептора 3 |
Экспрессия HIF1α в гемопоэтических стволовых клетках объясняет природу покоя стволовых клеток, поскольку они метаболически поддерживаются с низкой скоростью, чтобы сохранять эффективность стволовых клеток в течение длительных периодов жизненного цикла организма.
Сигнальный каскад HIF опосредует эффекты гипоксии, состояния низкой концентрации кислорода, на клетку. Гипоксия часто не дает клеткам дифференцироваться. Однако гипоксия способствует образованию кровеносных сосудов и важна для формирования сосудистой системы у эмбрионов и опухолей. Гипоксия в ранах также способствует миграции кератиноцитов и восстановлению эпителия. Поэтому неудивительно, что модуляция HIF-1 была определена как многообещающая парадигма лечения при заживлении ран.
В целом HIF жизненно важны для развития. У млекопитающих делеция генов HIF-1 приводит к перинатальной смерти. HIF-1 было показано, что жизненно важно для хондроцитов выживания, позволяя клеткам адаптироваться к условиям с низким содержанием кислорода в пределах роста пластин из костей. HIF играет центральную роль в регуляции метаболизма человека.
 Нобелевская премия по физиологии и медицине 2019 г.: Как клетки воспринимают кислород и адаптируются к нему. В нормоксических условиях Hif-1 альфа гидроксилируется по двум остаткам пролина. Затем он связывается с VHL и маркируется убиквитином, что приводит к протеасомной деградации. В условиях гипоксии Hif-1 альфа перемещается в ядро клетки и связывается с Hif-1 бета. Затем этот комплекс связывается с областью HRE ДНК, что приводит к транскрипции генов, которые участвуют во множестве процессов, включая эритропоэз, гликолиз и ангиогенез.
Нобелевская премия по физиологии и медицине 2019 г.: Как клетки воспринимают кислород и адаптируются к нему. В нормоксических условиях Hif-1 альфа гидроксилируется по двум остаткам пролина. Затем он связывается с VHL и маркируется убиквитином, что приводит к протеасомной деградации. В условиях гипоксии Hif-1 альфа перемещается в ядро клетки и связывается с Hif-1 бета. Затем этот комплекс связывается с областью HRE ДНК, что приводит к транскрипции генов, которые участвуют во множестве процессов, включая эритропоэз, гликолиз и ангиогенез. Альфа - субъединица HIF является гидроксилированными в консервативном пролине остатков по HIF пролил-гидроксилазу, что позволяет их признание и убиквитинирование со стороны VHL E3 лигазы убиквитина, который помечает их для быстрой деградации по протеасоме. Это происходит только в нормоксических условиях. В условиях гипоксии пролилгидроксилаза HIF ингибируется, поскольку она использует кислород в качестве косубстрата.
Ингибирование переноса электронов в комплексе сукцинатдегидрогеназы из-за мутаций в генах SDHB или SDHD может вызвать накопление сукцината, который ингибирует пролилгидроксилазу HIF, стабилизируя HIF-1α. Это называется псевдогипоксией.
HIF-1, когда он стабилизируется в условиях гипоксии, активирует несколько генов, чтобы способствовать выживанию в условиях низкого содержания кислорода. К ним относятся ферменты гликолиза, которые позволяют синтезировать АТФ кислородно-независимым образом, и фактор роста эндотелия сосудов (VEGF), который способствует ангиогенезу. HIF-1 действует путем связывания с гипоксически-чувствительными элементами (HRE) в промоторах, которые содержат последовательность NCGTG (где N представляет собой либо A, либо G). Последние работы из лабораторий Sonia Роча и Уильям Келин младший показывает, что гипоксия модулирует гистонов метилирование и перепрограммирует хроматином Эта статья была опубликована спина к спине с тем, что в 2019 Нобелевской премии по физиологии и медицине победителя для медицины Уильям Келин младший Эта работа был выделен в независимой редакционной статье.
Было показано, что мышечный A-киназа-заякоренный белок (mAKAP) организовывает E3 ubiquitin ligases, влияя на стабильность и позиционирование HIF-1 внутри сайта его действия в ядре. Истощение mAKAP или нарушение его нацеливания на перинуклеарную (в кардиомиоцитах) область изменяет стабильность HIF-1 и активацию транскрипции генов, связанных с гипоксией. Таким образом, «компартментализация» чувствительных к кислороду сигнальных компонентов может влиять на гипоксический ответ.
Расширенные знания о молекулярных механизмах регуляции активности HIF1 в условиях гипоксии резко контрастируют с недостатком информации о механистических и функциональных аспектах, управляющих опосредованной NF-κB регуляцией HIF1 в условиях нормоксии. Однако стабилизация HIF-1α также обнаруживается в негипоксических условиях посредством неизвестного до недавнего времени механизма. Было показано, что NF-κB (ядерный фактор κB) является прямым модулятором экспрессии HIF-1α в присутствии нормального давления кислорода. Исследования siRNA (малой интерферирующей РНК) для отдельных членов NF-κB выявили различное влияние на уровни мРНК HIF-1α, что указывает на то, что NF-κB может регулировать базальную экспрессию HIF-1α. Наконец, было показано, что, когда эндогенный NF-κB индуцируется обработкой TNFα (фактор некроза опухоли α), уровни HIF-1α также изменяются NF-κB-зависимым образом. HIF-1 и HIF-2 имеют разные физиологические роли. HIF-2 регулирует выработку эритропоэтина во взрослой жизни.
В нормальных условиях после травмы HIF-1a деградируют под пролилгидроксилазой гидроксилазы (PhDs). В июне 2015 года ученые обнаружили, что продолжающаяся повышающая регуляция HIF-1a с помощью ингибиторов PHD восстанавливает потерянные или поврежденные ткани у млекопитающих, у которых есть репаративный ответ; и продолжающееся подавление Hif-1a приводит к заживлению с ответом рубцевания у млекопитающих с предыдущим регенеративным ответом на потерю ткани. Акт регулирования HIF-1a может либо выключить, либо включить ключевой процесс регенерации млекопитающих. Одним из таких регенеративных процессов, в которых участвует HIF1A, является заживление кожи. Исследователи из Медицинской школы Стэнфордского университета продемонстрировали, что активация HIF1A способна предотвращать и лечить хронические раны у мышей с диабетом и старых мышей. Мало того, что раны у мышей зажили быстрее, но и качество новой кожи было даже лучше, чем у оригинала. Кроме того, был описан регенерирующий эффект модуляции HIF-1A на стареющие клетки кожи и продемонстрировано омолаживающее действие на стареющую кожу лица у пациентов. Модуляция HIF также была связана с благотворным влиянием на выпадение волос. Биотехнологическая компания Tomorrowlabs GmbH, основанная в Вене в 2016 году врачом Домиником Душером и фармакологом Домиником Тором, использует этот механизм. На основе активного ингредиента HSF («укрепляющий фактор HIF»), заявленного на получение патента, были разработаны продукты, которые должны способствовать регенерации кожи и волос.
Недавно было разработано несколько лекарств, которые действуют как селективные ингибиторы пролилгидроксилазы HIF. Наиболее известные соединения: роксадустат (FG-4592); Вададустат (AKB-6548), Дапродустат (GSK1278863), Дезидустат (ZYAN-1) и Молидустат (Bay 85-3934), все из которых предназначены в качестве пероральных препаратов для лечения анемии. Другие важные соединения из этого семейства, которые используются в исследованиях, но не были разработаны для медицинского применения на людях, включают MK-8617, YC-1, IOX-2, 2-метоксиэстрадиол, GN-44028, AKB-4924, Bay 87. -2243, FG-2216 и FG-4497. За счет ингибирования фермента пролилгидроксилазы стабильность HIF-2α в почках увеличивается, что приводит к увеличению эндогенной продукции эритропоэтина. Оба соединения FibroGen прошли в фазу II клинических испытаний, но они были временно приостановлены в мае 2007 года после смерти участника испытания, принимавшего FG-2216 от фульминантного гепатита (печеночная недостаточность), однако неясно, действительно ли эта смерть была вызвана FG-2216. Приостановление дальнейших испытаний FG-4592 было снято в начале 2008 г. после того, как FDA рассмотрело и одобрило подробный ответ FibroGen. Роксадустат, вададустат, дапродустат и молидустат в настоящее время прошли клинические испытания фазы III для лечения почечной анемии.
В других сценариях и в отличие от терапии, описанной выше, недавние исследования показывают, что индукция HIF при нормоксии может иметь серьезные последствия в условиях болезни с хроническим воспалительным компонентом. Также было показано, что хроническое воспаление является самовоспроизводящимся и искажает микросреду в результате аберрантно активных факторов транскрипции. Как следствие, изменения фактора роста, хемокинов, цитокинов и баланса ROS происходят в клеточной среде, которая, в свою очередь, обеспечивает ось роста и выживания, необходимую для развития de novo рака и метастазов. Эти результаты имеют многочисленные последствия для ряда патологий, при которых нарушена регуляция NF-κB и HIF-1, включая ревматоидный артрит и рак. Следовательно, считается, что понимание перекрестной связи между этими двумя ключевыми факторами транскрипции, NF-κB и HIF, значительно ускорит процесс разработки лекарств.
Активность HIF участвуют в ангиогенезе, необходимый для роста опухоли рака, поэтому ингибиторы HIF, таких как фенэтилизотиоцианат и акрифлавин являются (с 2006 г.) под следствием эффектов противораковых.
Исследования, проведенные на мышах, показывают, что стабилизация HIF с помощью ингибитора пролилгидроксилазы HIF улучшает память гиппокампа, вероятно, за счет увеличения экспрессии эритропоэтина. Активаторы пути HIF, такие как ML-228, могут обладать нейропротекторным действием и представляют интерес в качестве потенциальных средств лечения инсульта и травм спинного мозга.
Белзутифан является ингибитором индуцируемого гипоксией фактора-2α, который исследуется для лечения почечно-клеточного рака, ассоциированного с болезнью фон Хиппеля – Линдау.
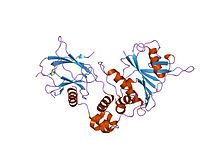 Структура комплекса HIF-1a-pVHL-ElonginB-ElonginC.
Структура комплекса HIF-1a-pVHL-ElonginB-ElonginC.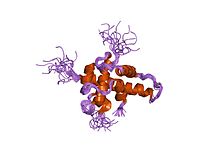 Структура альфа-субъединицы фактора-1, индуцируемого гипоксией.
Структура альфа-субъединицы фактора-1, индуцируемого гипоксией.