
Войти
| Polyomaviridae | |
|---|---|
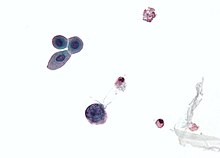 | |
| Микрофотография, показывающая инфицированную полиомавирусом клетку - большая (синяя) ячейка ниже центра -осталось. Цитологический образец мочи. | |
| Классификация вируса | |
| (без рейтинга): | Вирус |
| Область: | Моноднавирия |
| Королевство: | Шотокувиры |
| Тип: | Cossaviricota |
| Класс: | Papovaviricetes |
| Порядок: | Sepolyvirales |
| Семейство: | Polyomaviridae |
| Род | |
Polyomaviridae - это семейство вирусов, естественные Хозяева - это прежде всего млекопитающие и птицы. По состоянию на 2019 год насчитывается четыре признанных рода и 102 вида, девять из которых не относятся к роду. Известно, что 14 видов заражают людей, в то время как другие, такие как обезьяний вирус 40, были идентифицированы у людей в меньшей степени. Большинство этих вирусов очень распространены и обычно бессимптомны в большинстве изученных популяций людей. Вирус BK связан с нефропатией у почечного трансплантата и у пациентов с непереносимым твердым органом, вирусом JC с прогрессирующей мультифокальной лейкоэнцефалопатией и вирусом клеток Меркеля. с раком из клеток Меркеля.
 Визуализация икосаэдрического вирусного капсида, содержащего 72 пентамера полиомавируса мыши VP1, окрашены таким образом, что участки поверхности, расположенные ближе к внутреннему центру, выглядят синими, а более удаленные - красными. Получено из PDB : 1SIE .
Визуализация икосаэдрического вирусного капсида, содержащего 72 пентамера полиомавируса мыши VP1, окрашены таким образом, что участки поверхности, расположенные ближе к внутреннему центру, выглядят синими, а более удаленные - красными. Получено из PDB : 1SIE .Полиомавирусы - это необолочечные вирусы с двухцепочечной ДНК с круговыми геномами примерно 5000 пар оснований. Геном упакован в вирусный капсид диаметром примерно 40-50 нанометров, который имеет форму икосаэдра (симметрия Т = 7). Капсид состоит из 72 пентамерных капсомеров белка, называемого VP1, который способен к самосборке в замкнутый икосаэдр; каждый пентамер VP1 связан с одной молекулой одного из двух других капсидных белков, VP2 или VP3.
 Структура генома вируса WU, полиомавируса человека. Ранняя область показана слева и содержит белки TAg (опухолевый антиген); поздняя область находится справа и содержит белки капсида.
Структура генома вируса WU, полиомавируса человека. Ранняя область показана слева и содержит белки TAg (опухолевый антиген); поздняя область находится справа и содержит белки капсида. Геном типичного полиомавируса кодирует от 5 до 9 белков, разделенных на две транскрипционные области, называемые ранними и поздние регионы из-за времени во время заражения, в котором они транскрибируются. Каждая область транскрибируется РНК-полимеразой II клетки-хозяина в виде единственной пре-мессенджерской РНК, содержащей несколько генов. Ранняя область обычно кодирует два белка, малый и большой опухолевые антигены, продуцируемые альтернативным сплайсингом. Поздняя область содержит три структурных белка капсида VP1, VP2 и VP3, продуцируемые альтернативными стартовыми сайтами трансляции. Дополнительные гены и другие вариации на эту тему присутствуют в некоторых вирусах: например, полиомавирусы грызунов содержат третий белок, называемый средним опухолевым антигеном в ранней области, который чрезвычайно эффективен для индукции клеточной трансформации ; SV40 имеет дополнительный капсидный белок VP4; в некоторых примерах есть дополнительный регуляторный белок, называемый агнопротеином, экспрессируемый из поздней области. Геном также содержит некодирующую контрольную или регуляторную область, содержащую промоторы ранних и поздних областей, сайты старта транскрипции и точку начала репликации.
| Genus | Структура | Симметрия | Капсид | Геномное расположение | Геномная сегментация |
|---|---|---|---|---|---|
| Полиомавирус | Икосаэдрический | T=7 | Необолочечный | Круглый | Монодостаточный |
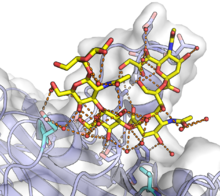 Полиомавирус мыши VP1 в комплексе с GT1a гликан. GT1a показан желтым, а мономер VP1 - с белой поверхностью и синим белковым скелетом. Сложная сеть водородных связей, многие из которых опосредованы водой, показана на поверхности связывания оранжевыми линиями, а участвующие белковые остатки показаны в виде полосок. Мутации двух остатков, выделенных голубым цветом в нижней части рисунка, могут существенно повлиять на патогенность. Из PDB : 5CPW .
Полиомавирус мыши VP1 в комплексе с GT1a гликан. GT1a показан желтым, а мономер VP1 - с белой поверхностью и синим белковым скелетом. Сложная сеть водородных связей, многие из которых опосредованы водой, показана на поверхности связывания оранжевыми линиями, а участвующие белковые остатки показаны в виде полосок. Мутации двух остатков, выделенных голубым цветом в нижней части рисунка, могут существенно повлиять на патогенность. Из PDB : 5CPW .Жизненный цикл полиомавируса начинается с проникновения в клетку-хозяин. Клеточные рецепторы полиомавирусов представляют собой остатки сиаловой кислоты гликанов, обычно ганглиозидов. Присоединение полиомавирусов к клеткам-хозяевам опосредуется связыванием VP1 с сиалированными гликанами на поверхности клетки. В некоторых конкретных вирусах происходят дополнительные взаимодействия клеточной поверхности; например, считается, что вирус JC требует взаимодействия с рецептором 5HT2A и вирусом клеток Меркеля с гепарансульфатом. Однако в целом взаимодействия вирус-клетка опосредуются обычно встречающимися молекулами на поверхности клетки и поэтому, вероятно, не являются основным фактором наблюдаемого клеточного типа тропизма отдельных вирусов. После связывания с молекулами на поверхности клетки вирион эндоцитозируется и попадает в эндоплазматический ретикулум - поведение, уникальное среди известных вирусов без оболочки, - где структура вирусного капсида может быть нарушается действием ферментов клетки-хозяина дисульфидизомеразы.
Детали перехода к ядру не ясны и могут варьироваться в зависимости от индивидуальных полиомавирусов. Часто сообщалось, что интактная, хотя и искаженная, частица вириона высвобождается из эндоплазматического ретикулума в цитоплазму клетки, где геном высвобождается из капсида, возможно, из-за низкой концентрации кальция в цитоплазме.. Как экспрессия вирусных генов, так и репликация вирусного генома происходят в ядре с использованием аппарата клетки-хозяина. Ранние гены, содержащие как минимум антиген малой опухоли (ST) и антиген большой опухоли (LT), экспрессируются первыми из одного , альтернативно сплайсированного информационная РНК цепь. Эти белки служат для управления клеточным циклом хозяина - нарушением регуляции перехода от фазы G1 к фазе S, когда геном клетки-хозяина реплицируется - потому что ДНК клетки-хозяина Репликационный аппарат необходим для репликации вирусного генома. Точный механизм этого нарушения регуляции зависит от вируса; например, SV40 LT может напрямую связываться с клеткой-хозяином p53, а мышиный полиомавирус LT - нет. LT индуцирует репликацию ДНК из некодирующей контрольной области вирусного генома (NCCR), после чего экспрессия ранней мРНК снижается и начинается экспрессия поздней мРНК, которая кодирует белки вирусного капсида. Когда эти взаимодействия начинаются, LT, принадлежащие нескольким полиомавирусам, включая полиомавирус клеток Меркеля, проявляют онкогенный потенциал. Было описано несколько механизмов регуляции перехода от ранней к поздней экспрессии генов, включая участие белка LT в репрессии раннего промотора, экспрессию незавершенных поздних мРНК с расширениями, комплементарными ранней мРНК, и экспрессию регуляторных микроРНК. Экспрессия поздних генов приводит к накоплению белков вирусного капсида в цитоплазме клетки-хозяина. Компоненты капсида проникают в ядро для инкапсидации новой вирусной геномной ДНК. Новые вирионы могут быть собраны на вирусных фабриках. Механизм высвобождения вируса из клетки-хозяина варьируется среди полиомавирусов; некоторые экспрессируют белки, которые облегчают выход из клетки, такие как агнопротеин или VP4. В некоторых случаях высокие уровни инкапсидированного вируса приводят к лизису клеток с высвобождением вирионов.
| Род | Сведения о хозяине | Тропизм ткани | Вход подробности | Сведения о выпуске | Сайт репликации | Сайт сборки | Передача |
|---|---|---|---|---|---|---|---|
| Полиомавирус | Млекопитающие; птицы | Дыхательная система; почки, мозг | эндоцитоз клеточного рецептора | Лизис | ядро | ядро | орально-фекальный |
антиген большой опухоли играет ключевую роль в регуляции жизненного цикла вируса, связываясь с вирусным источником репликации ДНК, где он способствует синтезу ДНК. Также, поскольку полиомавирус полагается на механизм клетки-хозяина для репликации, клетка-хозяин должна находиться в s-фазе, чтобы это началось. Благодаря этому большой Т-антиген также модулирует клеточные сигнальные пути, чтобы стимулировать развитие клеточного цикла, связываясь с рядом клеточных контрольных белков. Это достигается с помощью двухсторонней атаки: ингибирования генов, подавляющих опухоль, p53 и членов семейства ретинобластомы (pRB), и стимуляции путей роста клеток путем связывания клеточной ДНК, АТФаза-геликаза, ассоциации ДНК-полимеразы α и связывание факторов преинициации транскрипции. Эта ненормальная стимуляция клеточного цикла - мощная сила для онкогенной трансформации.
Белок антиген малой опухоли также способен активировать несколько клеточных путей, которые стимулируют пролиферацию клеток. Малые Т-антигены полиомавируса обычно нацелены на протеинфосфатазу 2A (PP2A ), ключевой мультисубъединичный регулятор множества путей, включая Akt, путь митоген-активируемой протеинкиназы (MAPK) и стресс- путь активированной протеинкиназы (SAPK). полиомавирус клеток Меркеля малый Т-антиген кодирует уникальный домен, называемый LT-стабилизационным доменом (LSD), который связывается и ингибирует FBXW7 E3-лигаза, регулирующая как клеточные, так и вирусные онкопротеины. В отличие от SV40, малый Т-антиген MCV непосредственно трансформирует клетки грызунов in vitro.
средний опухолевый антиген используется в модельных организмах, разработанных для изучения рака, таких как система MMTV-PyMT, где средний T соединен с промотором MMTV . Там он функционирует как онкоген, а ткань, в которой развивается опухоль, определяется промотором MMTV.
Капсид полиомавируса состоит из одного основного компонента, основного капсидного белка VP1 и одного или двух второстепенных компонентов, второстепенных капсидных белков VP2 и VP3. Пентамеры VP1 образуют закрытый икосаэдрический вирусный капсид, и внутри капсида каждый пентамер связан с одной молекулой VP2 или VP3. Некоторые полиомавирусы, такие как полиомавирус клеток Меркеля, не кодируют и не экспрессируют VP3. Белки капсида экспрессируются из поздней области генома.
агнопротеин представляет собой небольшой многофункциональный фосфопротеин, обнаруженный в поздней кодирующей части генома. некоторых полиомавирусов, в первую очередь вируса BK, вируса JC и SV40. Он важен для пролиферации вирусов, которые его экспрессируют, и считается, что он участвует в регуляции жизненного цикла вируса, особенно репликации и выхода вируса из клетки-хозяина, но точные механизмы неясны.
Полиомавирусы относятся к группе I (вирусы дцДНК). Классификация полиомавирусов была предметом нескольких предложенных изменений по мере обнаружения новых членов группы. Раньше полиомавирусы и папилломавирусы, которые имеют много общих структурных особенностей, но очень разные геномные организации, были объединены в уже устаревшее семейство Papovaviridae. (Название Papovaviridae образовано от трех сокращений: Pa для папилломавируса, Po для полиомавируса и Va для «вакуолизации».) Полиомавирусы были разделены на три основные клады (то есть генетически связанные группы): Клады SV40, клады птиц и клады полиомавирусов мышей. В последующей предложенной реклассификации Международного комитета по таксономии вирусов (ICTV) было рекомендовано разделить семейство Polyomaviridae на три рода:
Текущая классификационная система ICTV распознает четыре рода и 102 вида, из которых девять не могут быть отнесены к роду. Эта система сохраняет различие между вирусами птиц и млекопитающих, группируя птичью субпопуляцию в род Gammapolyomavirus.
Следующие виды не отнесены к роду:
Описание дополнительных вирусов продолжается. включают полиомавирус калана 1 и полиомавирус альпаки A Другой вирус - полиомавирус гигантской панды 1. Другой вирус был описан от грызунов-сигмодонтинов. Другой - полиомавирус 1 бурозубки - был описан для бурозубки.
Большинство полиомавирусов не заражают людей. Из полиомавирусов, каталогизированных по состоянию на 2017 год, в общей сложности 14 были известны с человеческими хозяевами. Однако некоторые полиомавирусы связаны с заболеванием человека, особенно у лиц с ослабленным иммунитетом. MCV сильно отличается от других полиомавирусов человека и наиболее близок к полиомавирусу мышей. Полиомавирус, связанный с Trichodysplasia spinulosa (TSV), отдаленно родственен MCV. Два вируса - HPyV6 и HPyV7 - наиболее тесно связаны с вирусами KI и WU, в то время как HPyV9 наиболее тесно связан с лимфотропным полиомавирусом африканских зеленых обезьян (LPV).
Описан четырнадцатый вирус. Полиомавирус Lyon IARC относится к полиомавирусу енота.
Были идентифицированы следующие 14 полиомавирусов с человеческими хозяевами, и их геномы секвенированы по состоянию на 2017 г.:
| Виды | Предлагаемый род | Название вируса | Аббревиатура | NCBI RefSeq | Год открытия | Клинический коррелят (если есть) | Список литературы |
|---|---|---|---|---|---|---|---|
| Полиомавирус человека 5 | Альфа | Полиомавирус клеток Меркеля | MCPyV | NC_010277 | 2008 | Рак из клеток Меркеля | |
| Полиомавирус человека 8 | Альфа | Полиомавирус Trichodysplasia spinulosa | TSPyV | NC_014361 | 2010 | Trichodysplasia spinulosa | |
| Полиомавирус человека 9 | Альфа | Полиомавирус человека 9 | HP | NC_015150 | 2011 | Неизвестно | |
| Полиомавирус человека 12 | Альфа | Полиомавирус человека 12 | HPyV12 | NC_020890 | 2013 | Неизвестно | |
| Полиомавирус человека 13 | Альфа | Полиомавирус Нью-Джерси | NJPyV | NC_024118 | 2014 | Неизвестно | |
| Полиомавирус человека 1 | Бета | BK полиомавирус | BKPyV | NC_001538 | 1971 | Полиомавирус-ассоциированная нефропатия ; геморрагический цистит | |
| полиомавирус человека 2 | бета | полиомавирус JC | JCPyV | NC_001699 | 1971 | Прогрессирующая мультифокальная лейкоэнцефалопатия | |
| полиомавирус человека 3 | Бета | полиомавирус KI | KIPyV | NC_009238 | 2007 | Неизвестно | |
| полиомавирус человека 4 | Бета | полиомавирус WU | WUPyV | NC_009539 | 2007 | Неизвестно | |
| Полиомавирус человека 6 | Дельта | Полиомавирус человека 6 | HPyV6 | NC_014406 | 2010 | Зудящий и дискератотический дерматоз, ассоциированный с HPyV6 (H6PD) | |
| Полиомавирус человека 7 | Дельта | Полиомавирус человека 7 | HPyV7 | NC_014407 | 2010 | Эпителиальная гиперплазия, связанная с HPyV7 | |
| Полиомавирус человека 10 | Дельта | Полиомавирус MW | MWPyV | NC_018102 | 2012 | Неизвестно | |
| Полиомавирус человека 11 | Дельта | Полиомавирус STL | STLPyV | NC_020106 | 2013 | Неизвестно | |
| Полиомавирус человека 14 | Альфа | Полиомавирус Лиона IARC | LIPyV | NC_034253.1 | 2017 | Неизвестно |
Дельтаполиомавирус содержит только четыре вируса человека, представленных в таблице выше, с типовым видом полиомавируса человека 6. Группы альфа и бета содержат вирусы, поражающие различных млекопитающих. Группа «Гамма» содержит вирусы птиц. Клинически значимые ассоциации заболеваний показаны только там, где ожидается причинная связь.
Антитела к лимфотропному полиомавирусу обезьян были обнаружены у людей, что позволяет предположить, что этот вирус - или близкородственный вирус - может инфицировать людей.
Все полиомавирусы являются очень распространенными инфекциями у детей и молодых взрослых. Большинство из этих инфекций практически не вызывают симптомов. Эти вирусы, вероятно, сохраняются на протяжении всей жизни почти у всех взрослых. Заболевания, вызываемые полиомавирусными инфекциями человека, наиболее распространены среди людей с ослабленным иммунитетом ; ассоциации заболевания включают вирус BK с нефропатией у почечного трансплантата и пациентов с непереносимым твердым органом, вирус JC с прогрессирующей мультифокальная лейкоэнцефалопатия и вирус клеток Меркеля (MCV) с раком из клеток Меркеля.
SV40 реплицируется в почках обезьян без вызывает заболевание, но может вызвать рак у грызунов в лабораторных условиях. В 1950-х и начале 1960-х годов более 100 миллионов человек могли подвергнуться воздействию SV40 из-за ранее не обнаруженного заражения SV40 вакциной от полиомиелита, что вызвало обеспокоенность по поводу возможности того, что вирус может вызывать заболевание у людей. Хотя сообщалось, что он присутствует при некоторых раковых заболеваниях человека, включая опухоли головного мозга, опухоли костей, мезотелиомы и неходжкинские лимфомы точное обнаружение часто затрудняется высоким уровнем перекрестной реактивности SV40 с широко распространенными полиомавирусами человека. Большинство вирусологов отвергают SV40 как причину рака человека.
Диагноз полиомавируса почти всегда ставится после первичной инфекции, поскольку он протекает бессимптомно или субклинически. Анализы антител обычно используются для определения наличия антител против отдельных вирусов. Чтобы отличить очень похожие полиомавирусы, часто необходимы конкурентные анализы.
В случаях прогрессирующей мультифокальной лейкоэнцефалопатии (ПМЛ) перекрестно-реактивные антитела к Т-антигену SV40 (обычно Pab419) используются для непосредственного окрашивания тканей на наличие антигена Т вируса JC. ПЦР можно использовать при биопсии ткани или спинномозговой жидкости для амплификации ДНК полиомавируса. Это позволяет не только обнаруживать полиомавирус, но и определять его подтип.
Для диагностики реактивации полиомавируса при полиомавирусной нефропатии (ПВН) используются три основных диагностических метода: цитология мочи, количественная оценка вирусная нагрузка как в моче, так и в крови, и биопсия почек. Реактивация полиомавируса в почках и мочевыводящих путях вызывает выделение инфицированных клеток, вирионов и / или вирусных белков с мочой. Это позволяет цитологическому исследованию мочи исследовать эти клетки, что в случае включения ядра полиомавируса является диагностическим признаком инфекции. Кроме того, поскольку моча инфицированного человека будет содержать вирионы и / или вирусную ДНК, количественное определение вирусной нагрузки может быть выполнено с помощью ПЦР. То же самое и с кровью.
Биопсия почек также может использоваться, если два только что описанных метода не позволяют сделать окончательные выводы или если требуется определенная вирусная нагрузка на почечную ткань. Подобно цитологии мочи, почечные клетки исследуются под световой микроскопией на наличие полиомавируса в ядре, а также на лизис клеток и вирусные частицы во внеклеточной жидкости. Вирусную нагрузку, как и раньше, также измеряют с помощью ПЦР.
Окрашивание тканей с использованием моноклональных антител против Т-антигена MCV показывает полезность при дифференциации карциномы из клеток Меркеля от других небольших круглоклеточных опухолей. Были разработаны анализы крови для обнаружения антител к MCV, которые показывают, что инфицирование вирусом широко распространено, хотя пациенты с карциномой из клеток Меркеля имеют исключительно более высокие ответы антител, чем лица, инфицированные бессимптомно.
Вирус JC является многообещающим генетическим маркером эволюции и миграции человека. Он переносится 70–90 процентами людей и обычно передается от родителей к потомству. Этот метод не представляется надежным для отслеживания недавнего африканского происхождения современных людей.
Полиомавирус мыши был первым обнаруженным полиомавирусом, о котором сообщил Людвик Гросс в 1953 г. как экстракт мышей лейкозов, способный вызывать опухоли околоушной железы. Возбудитель был идентифицирован как вирус Сарой Стюарт и Бернис Эдди, в честь которых он когда-то был назван «полиома SE». Термин «полиома» относится к способности вирусов продуцировать множественные (поли-) опухоли (-омы) при определенных условиях. Название критиковалось как «лингвистический бутерброд без мяса» («без мяса», потому что обе морфемы в слове «полиома» являются аффиксами), дающий мало информации о биологии вирусов; на самом деле, последующие исследования показали, что большинство полиомавирусов редко вызывают клинически значимые заболевания у их организмов-хозяев в естественных условиях.
По состоянию на 2017 г. были идентифицированы и секвенированы десятки полиомавирусов, заражающих в основном птиц и млекопитающих. Известно, что два полиомавируса поражают рыб: черный морской окунь и морской лещ. Всего известно четырнадцать полиомавирусов, инфицирующих людей.