
Войти
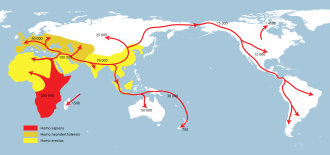 Последовательное расселение. Homo erectus наибольшее распространение (желтый),. Homo neanderthalensis наибольшее распространение (охра) и. Homo sapiens (красный цвет).
Последовательное расселение. Homo erectus наибольшее распространение (желтый),. Homo neanderthalensis наибольшее распространение (охра) и. Homo sapiens (красный цвет).  Распространение ранних современных людей из Африки через Ближний Восток
Распространение ранних современных людей из Африки через Ближний Восток  Карта миграции современного человека из Африки, основанная на митохондриальной ДНК. Цветные кольца указывают на тысячу лет до настоящего.
Карта миграции современного человека из Африки, основанная на митохондриальной ДНК. Цветные кольца указывают на тысячу лет до настоящего. В палеоантропологии, недавнее африканское происхождение современных людей, также называемых «Out of Africa » теория (OOA ), недавняя гипотеза единственного происхождения (RSOH ), гипотеза замены или недавняя Модель африканского происхождения (RAO ) является доминирующей моделью географического происхождения и ранней миграции анатомически современных людей (Homo sapiens ). Она следует за ранним распространением гомининов из Африки, осуществленным Homo erectus, а затем Homo neanderthalensis.
Модель предлагает «единое происхождение» Homo sapiens в таксономическом смысле, что исключает параллельную эволюцию признаков, считающихся анатомически современными в других регионах, но не исключает множественное смешение между H. sapiens и архаичным люди в Европе и Азии. H. sapiens, скорее всего, возник в Африканском Роге между 300 000 и 200 000 лет назад. Модель «недавнего африканского происхождения» предполагает, что все современные неафриканские популяции в значительной степени произошли от популяций H. sapiens, покинувших Африку после этого времени.
Было по крайней мере несколько расселений современных людей "из Африки", возможно, начавшееся уже 270 000 лет назад, в том числе 215 000 лет назад по крайней мере до Греции и, конечно, через Северная Африка около 130 000–115 000 лет назад. Эти ранние волны, по-видимому, в основном вымерли или отступили к 80 000 лет назад.
Наиболее значительная «недавняя» волна произошла около 70 000–50 000 лет назад по так называемому «Южному маршруту ", быстро распространившись вдоль побережья Азии и достигнув Австралии примерно 65 000–50 000 лет назад (хотя некоторые исследователи ставят под сомнение более ранние австралийские даты и считают, что люди прибыли туда не ранее 50 000 лет назад)., в то время как другие предположили, что эти первые поселенцы Австралии могут представлять более старую волну до более значительной миграции из Африки и, следовательно, не обязательно быть предками более поздних жителей региона), в то время как Европа была заселена ранним ответвлением, которое заселило Ближний Восток и Европа менее 55000 лет назад.
В 2010-х годах исследования популяционной генетики обнаружили доказательства скрещивания, которое имело место между H. sapiens и архаическими людьми в Евразии, Океании и Африке, указывая t Современные группы населения, хотя и происходят от ранних H. sapiens, в меньшей степени также происходят от региональных вариантов архаичных людей.
 последовательность слоев в Ксар Акил в левантийском коридоре и обнаружение двух окаменелостей Homo sapiens, датировано 40 800 т. o 39 200 лет назад для "Egbert" и 42 400–41 700 лет назад для "Ethelruda".
последовательность слоев в Ксар Акил в левантийском коридоре и обнаружение двух окаменелостей Homo sapiens, датировано 40 800 т. o 39 200 лет назад для "Egbert" и 42 400–41 700 лет назад для "Ethelruda". "Недавнее африканское происхождение" или Out of Africa II относится к миграции анатомически современных людей (Homo sapiens ) из Африки после их появления в ок. 300 000–200 000 лет назад, в отличие от «Out of Africa I », в котором говорится о миграции архаичных людей из Африки в Евразию примерно 1,8–0,5 миллиона лет назад. Омо-Кибиш I (Омо I) из южной Эфиопии - самый старый из известных в настоящее время анатомически современных скелетов Homo sapiens (196 ± 5 тыс. Лет назад).
С начала 21 века картина «недавнего единственного происхождения» "миграции стали значительно более сложными не только из-за открытия примеси современности и архаики, но и из-за растущего числа свидетельств того, что" недавняя миграция из Африки "происходила в виде нескольких волн, распространявшихся в течение длительного периода времени. По состоянию на 2010 г. существовало два основных принятых пути расселения из Африки ранних анатомически современных людей: через «Северный путь» (через долину Нила и Синай) и «Южный путь» через Баб al Mandab пролив.
 Анатомически современные люди, известные археологические останки в Европе и Африке, прямо датированные, откалиброванные по углеродным датам на 2013.
Анатомически современные люди, известные археологические останки в Европе и Африке, прямо датированные, откалиброванные по углеродным датам на 2013. Начиная с 135 000 лет назад, в тропической Африке происходили мегапроходы, которые вытеснили людей с суши к морским берегам и вынудили их перейти на другие континенты.
Современные люди пересек пролив Баб-эль-Мандеб в южной части Красного моря и двинулся вдоль зеленых берегов вокруг Аравии, а оттуда в остальную часть Евразии. Окаменелости ранних Homo sapiens были найдены в пещере Кафзех в Израиле и датированы от 80 000 до 100 000 лет назад. Похоже, что эти люди либо вымерли, либо отступили обратно в Африку 70 000–80 000 лет назад, возможно, их заменили движущиеся на юг неандертальцы, бежавшие из более холодных регионов Европы ледникового периода. Хуа Лю и др. проанализировали аутосомные микросателлитные маркеры, датируемые примерно 56 000 лет назад. Они интерпретируют палеонтологическую окаменелость как изолированное раннее ответвление, вернувшееся обратно в Африку.
Открытие каменных орудий в Объединенных Арабских Эмиратах в 2011 г. на Участок Фая-1 в Млейха, Шарджа, указывает на присутствие современных людей по крайней мере 125000 лет назад, что привело к возрождению "давно забытых" североафриканских
В Омане Бьен Джовен в 2011 году обнаружил участок, на котором находилось более 100 фрагментов каменных орудий, принадлежащих позднему Нубийскому комплексу, ранее известного только из археологических раскопок. раскопки в Судане. По двум оценкам возраста, полученным с помощью оптически стимулированной люминесценции, возраст арабско-нубийского комплекса составляет примерно 106 000 лет. Это свидетельствует о наличии особого технокомплекса каменного века на юге Аравии, около более ранней части морского изотопного этапа 5.
. Согласно Кульвилму и его соавторам, неандертальцы генетически способствовали современному человеку, живущему за пределами Африки около 100000 лет назад: люди, которые уже отделились от других современных людей около 200000 лет назад, и эта ранняя волна современных людей за пределами Африки также генетически внесла свой вклад в алтайские неандертальцы. Они обнаружили, что «предки неандертальцев с Горного Алтая и первые современные люди встретились и скрестились, возможно, на Ближнем Востоке, на много тысяч лет раньше, чем считалось ранее». По словам соавтора Илана Гронау, «это фактически дополняет археологические свидетельства присутствия ранних современных людей из Африки около 100 000 лет назад и раньше, предоставляя первые генетические свидетельства существования таких популяций». Подобные генетические примеси события были отмечены и в других регионах.
В Китае человек Люцзян (китайский >: 柳江 人) - один из первых современных людей, обитающих в Восточной Азии. Дата, которую чаще всего приписывают останкам, - 67000 лет назад. Высокие показатели изменчивости, полученные с помощью различных методик датирования, проведенных разными исследователями, помещают наиболее широко принятый диапазон дат как минимум в 67000 лет назад, но не исключают датировки до 159000 лет назад. Liu, Martinón-Torres et al. al. (2015) утверждают, что современные человеческие зубы были найдены в Китае не менее 80000 лет назад.
 Красное море пересечение
Красное море пересечение Примерно 50–70 000 лет назад часть носителей митохондриальной гаплогруппы L3 мигрировала из Восточной Африки на Ближний Восток. Было подсчитано, что из населения от 2000 до 5000 человек в Африке только небольшая группа, возможно, всего от 150 до 1000 человек, пересекла Красное море. Группа, которая пересекла Красное море, путешествовала по прибрежному маршруту вокруг Аравии и Персидского плато в Индию, которая, по-видимому, была первым крупным пунктом поселения. Уэллс (2003)) приводил доводы в пользу маршрута вдоль южного побережья Азии примерно через 250 километров (155 миль), достигнув Австралии около 50 000 лет назад.
Сегодня в Баб-эль-Мандебском проливе Красное море имеет ширину около 20 километров (12 миль), но 50 000 лет назад уровень моря составлял 70 метров. (230 футов) ниже (из-за оледенения), и вода была намного уже. Хотя проливы никогда не закрывались полностью, они были достаточно узкими, чтобы их можно было пересечь на простых плотах, и между ними могли быть острова. В Эритрее были найдены ракушки кучи 125000 лет, что указывает на то, что в рацион древних людей входили морепродукты, добытые пляжным прочесыванием.
Датировка Южного Расселения является вопросом спор. Это могло произойти либо до, либо после Тоба, катастрофического извержения вулкана, которое произошло между 69 000 и 77 000 лет назад на месте современного озера Тоба. Каменные орудия, обнаруженные под слоями пепла в Индии, могут указывать на расселение до Тоба, но источник этих орудий оспаривается. Признаком пост-Тоба является гаплогруппа L3, которая возникла до расселения людей из Африки и может быть датирована 60 000–70 000 лет назад, «предполагая, что человечество покинуло Африку через несколько тысяч лет после Тоба». Некоторые исследования, показывающие более медленные, чем ожидалось, генетические мутации в ДНК человека, были опубликованы в 2012 году, что указывает на пересмотренную датировку миграции между 90 000 и 130 000 лет назад. Некоторые недавние исследования предполагают миграцию из Африки около 50 000–65 000 лет назад предков современных неафриканских популяций, что аналогично большинству предыдущих оценок.
A окаменелость современного человека, датируемая 54 700 лет назад, была найдена в пещере Манот в Израиле, названной Манот 1, хотя Groucutt et al. (2015).
Считается, что Австралия была заселена примерно 65 000–50 000 лет назад. По состоянию на 2017 год самым ранним свидетельствам существования людей в Австралии не менее 65000 лет, в то время как Макчесни заявил, что
... генетические данные свидетельствуют о том, что небольшая группа с маркером M168 мигрировала из Африки вдоль побережья Аравийского полуострова. и Индия, через Индонезию, и достигла Австралии очень рано, между 60 000 и 50 000 лет назад. Эта очень ранняя миграция в Австралию также подтверждается Rasmussen et al. (2011).
Окаменелости из озера Мунго, Австралия, были датированы примерно 42000 лет назад. Другие окаменелости с места под названием Мадджедбебе были датированы по крайней мере 65 000 лет назад, хотя некоторые исследователи сомневаются в этой ранней оценке и датируют окаменелости Маджедбебе примерно 50 000 лет назад самым старым.
Тяньюаньский человек из Китая имеет вероятный диапазон дат между 38000 и 42000 лет назад, тогда как Люцзян из того же региона имеет вероятный диапазон дат между 67000 и 159000 лет назад. Согласно тестам ДНК 2013 года, человек из провинции Тяньюань является родственником «многих современных азиатов и коренных американцев ». Тяньюань по морфологии похож на человека Минатогава, современных людей, датируемых 17000-19000 лет назад и найденных на острове Окинава, Япония.
Согласно Macaulay et al. (2005), раннее ответвление от южного расселения с гаплогруппой N следовало по Нилу из Восточной Африки, направляясь на север и пересекая Азию через Синай. Затем эта группа разветвлялась: одни двинулись в Европу, а другие на восток, в Азию. Эта гипотеза подтверждается относительно поздней датой прибытия современных людей в Европу, а также археологическими данными и данными ДНК. На основании анализа 55 митохондриальных геномов человека (мтДНК) охотников-собирателей Posth et al. (2016) приводят доводы в пользу «быстрого однократного расселения всех неафриканцев менее 55 000 лет назад».
 Карта ранней диверсификации современного человека согласно митохондриальной популяционной генетике (см.: Гаплогруппа L ).
Карта ранней диверсификации современного человека согласно митохондриальной популяционной генетике (см.: Гаплогруппа L ).Первой ветвью, ответвленной от Митохондриальной Евы, была L0. Эта гаплогруппа широко распространена среди Сан в Южной Африке и сандаве в Восточной Африке. Он также встречается среди людей мбути. Эти группы разветвились в начале истории человечества и с тех пор оставались относительно генетически изолированными. Гаплогруппы L1, L2 и L3 являются потомками L1 – L6 и в основном ограничены Африкой. Макрогаплогруппы M и N, которые являются линиями остального мира за пределами Африки, происходят от L3. L3 около 70 000 лет, в то время как гаплогруппы M и N имеют возраст примерно 65-55 000 лет. Связь между такими деревьями генов и демографической историей все еще обсуждается, когда
Из всех линий, присутствующих в Африке, только потомки женского пола одной линии, гаплогруппы L3 мтДНК, находятся за пределами Африки. Если бы было несколько миграций, можно было бы ожидать, что будут найдены потомки более чем одной линии. Потомки женского пола L3, линии гаплогрупп M и N, встречаются в Африке с очень низкой частотой (хотя популяции гаплогруппы M1 очень древние и разнообразны в Северная и Северо-Восточная Африка ) и, по всей видимости, появились недавно. Возможное объяснение состоит в том, что эти мутации произошли в Восточной Африке незадолго до массового исхода и впоследствии стали доминирующими гаплогруппами посредством эффекта основателя. Альтернативно, мутации могли возникнуть вскоре после этого.
Результаты мтДНК, собранной у коренных малазийцев, называемых оранг асли, показывают, что гапологруппы M и N имеют общие характеристики с исходными африканскими группами примерно 85000 лет назад и имеют общие характеристики с субгаплогруппами, обнаруженными в прибрежных регионах Юго-Восточной Азии, таких как Австралазия, Индийский субконтинент и по всей континентальной Азии, которые рассеялись и отделились от своего африканского прародителя примерно 65000 лет назад. Это распространение на южном побережье могло произойти до распространения через Левант примерно 45 000 лет назад. Эта гипотеза пытается объяснить, почему гаплогруппа N преобладает в Европе и почему гаплогруппа M отсутствует в Европе. Считается, что свидетельства прибрежной миграции были уничтожены повышением уровня моря в эпоху голоцена. В качестве альтернативы, небольшая европейская популяция-основатель, которая сначала выражала гаплогруппу M и N, могла потерять гаплогруппу M из-за случайного генетического дрейфа в результате узкого места (то есть эффекта основателя ).
Группа, которая пересекла Красное море, путешествовала по прибрежному маршруту вокруг Аравии и Персии, пока не достигла Индии. Гаплогруппа M находится в высоких распространены вдоль южных прибрежных регионов Пакистана и Индии, и наибольшее разнообразие наблюдается в Индии, что указывает на то, что именно здесь, возможно, произошла мутация. Шестьдесят процентов населения Индии принадлежат к гаплогруппе M. Коренные жители Андаманских островов также принадлежат к роду М. Считается, что андаманцы являются ответвлениями некоторых из первых жителей Азии из-за их долгой изоляции от материка. Они свидетельствуют о прибрежном пути ранних поселенцев, который простирается от Индии до Таиланда и Индонезии до востока Новой Гвинеи. Поскольку M часто встречается у горцев из Новой Гвинеи, а андаманцы и жители Новой Гвинеи имеют темную кожу и афро-текстурированные волосы, некоторые ученые считают, что все они являются частью одной и той же волны мигрантов, уехавших через Красное море ~ 60 000 лет назад в Великой миграции прибрежных районов. Доля гаплогруппы M увеличивается на восток от Аравии до Индии; в восточной Индии M превосходит N в соотношении 3: 1. Пересекая Юго-Восточную Азию, гаплогруппа N (в основном в форме производных от ее субклада R) снова появляется как преобладающая линия. M преобладает в Восточной Азии, но среди коренных австралийцев N - более распространенная линия. Такое случайное распространение гаплогруппы N из Европы в Австралию можно объяснить эффектами основателя и узкими местами населения.
Исследование африканского, европейского и азиатского населения в 2002 г., обнаружили большее генетическое разнообразие среди африканцев, чем среди евразийцев, и что генетическое разнообразие среди евразийцев в значительной степени является подмножеством генетического разнообразия среди африканцев, что подтверждает модель «вне Африки». Большое исследование Coop et al. (2009) нашли доказательства естественного отбора в аутосомной ДНК за пределами Африки. В исследовании различаются неафриканские развертки (в частности, варианты KITLG, связанные с цветом кожи ), западно-евразийские развертки (SLC24A5 ) и восточно-азиатские развертки (MC1R, соответствует цвету кожи). На основании этих данных исследование пришло к выводу, что человеческие популяции столкнулись с новым селективным давлением по мере их распространения за пределы Африки. MC1R и его связь с цветом кожи уже обсуждались Liu, Harding et al. (2000), стр. 135 ошибка harvp: нет цели: CITEREFLiu, _Harding_et_al.2000 (help ). Согласно этому исследованию, жители Папуа-Новой Гвинеи продолжали подвергаться отбору по темному цвету кожи, так что, хотя эти группы отличаются от африканцев в других местах, аллель темного цвета кожи, характерный для современных африканцев, андаманцев и новогвинейцев, является архаизмом.. Эндикотт и др. (2003) предполагают конвергентную эволюцию. Исследование 2014 года, проведенное Гурдасани и соавт. указывает на то, что более высокое генетическое разнообразие в Африке было дополнительно увеличено в некоторых регионах в результате относительно недавних миграций из Евразии, затронувших некоторые части Африки.
Другой многообещающий путь к реконструкции генетической генеалогии человека - через вирус JC (JCV), тип человеческого полиомавируса, который переносится 70–90 процентами людей и который обычно передается вертикально от родителей к потомству, что свидетельствует о совместном расхождении с популяциями людей. По этой причине JCV использовался в качестве генетического маркера эволюции и миграции человека. Этот метод не представляется надежным для миграции из Африки, в отличие от генетики человека, штаммы JCV, связанные с африканскими популяциями, не являются базовыми. Из этого Shackelton et al. (2006) пришли к выводу, что либо базальный африканский штамм JCV вымер, либо первоначальная инфекция JCV возникла после миграции из Африки.
Доказательства существования архаичных человеческих видов (происходящих от Homo heidelbergensis ), имеющих скрещивание с современных людей за пределами Африки был обнаружен в 2010-х годах. В первую очередь это касается неандертальской примеси во всех современных популяциях, за исключением африканцев к югу от Сахары, но также были представлены доказательства примеси гоминина Денисова в Австралазии (т.е. у меланезийцев, австралийских аборигенов и некоторых негритосов ).
Скорость поступления примеси неандертальцев в европейское и азиатское население по состоянию на 2017 год оценивается примерно в 2–3%.
Архаическая примесь в некоторых группах охотников-собирателей африканского населения к югу от Сахары (биака пигмеи и сан ), происходящая от отделившихся архаичных гомининов от современного человеческого происхождения около 700 000 лет назад, был обнаружен в 2011 году. Уровень примеси оценивался примерно в 2%. Примесь из архаичных гомининов еще более раннего времени расхождения, по оценкам, от 1,2 до 1,3 миллиона лет назад, была обнаружена в Пигмеи, Хадза и пять Сандаве в 2012 году. Из анализа Муцина 7, сильно расходящегося гаплотипа th По оценкам, время слияния at с другими вариантами составляет около 4,5 миллионов лет назад, и, как считается, он является специфическим для африканских популяций и, как предполагается, был получен в результате скрещивания африканских современных и архаичных людей.
In Помимо генетического анализа, Petraglia et al. также исследуются небольшие каменные орудия труда (микролитические материалы) с индийского субконтинента и объясняется рост населения на основе реконструкции палеосреды. Он предположил, что каменные орудия в Южной Азии могут быть датированы 35 тыс. Лет назад, а на новую технологию могут повлиять изменения окружающей среды и давление населения.
 Фронтиспис книги Хаксли Доказательства места человека в природе (1863): изображение сравнивает скелет человека с другими обезьянами.
Фронтиспис книги Хаксли Доказательства места человека в природе (1863): изображение сравнивает скелет человека с другими обезьянами. Кладистические отношения людей с африканскими обезьянами был предложен Чарльзом Дарвином после изучения поведения африканских обезьян, одна из которых была выставлена в Лондонском зоопарке. Анатом Томас Хаксли также поддержал эту гипотезу и предположил, что африканские обезьяны имеют тесные эволюционные отношения с людьми. Этим взглядам противостоял немецкий биолог Эрнст Геккель, который был сторонником теории «Из Азии». Геккель утверждал, что люди более тесно связаны с приматами Юго-Восточной Азии, и отверг африканскую гипотезу Дарвина.
В Происхождении человека Дарвин предположил, что люди произошли от обезьян, которые по-прежнему обладали маленьким мозгом, но ходили прямо, освобождая руки для целей, способствующих разуму; он думал, что такие обезьяны были африканскими:
В каждом большом регионе мира живые млекопитающие тесно связаны с вымершими видами того же региона. Следовательно, вероятно, что в Африке раньше жили вымершие обезьяны, тесно связанные с гориллами и шимпанзе ; и поскольку эти два вида теперь являются ближайшими союзниками человека, несколько более вероятно, что наши ранние предки жили на африканском континенте, чем где-либо еще. Но спекулировать на эту тему бесполезно, поскольку обезьяна размером с человека почти такая же, как дриопитек из Лартета, который был тесно связан с антропоморфными гилобатами, существовавшими в Европе. в период верхнего миоцена ; и с тех пор как столь отдаленный период Земля, несомненно, претерпела множество великих революций, и было достаточно времени для миграции в самых больших масштабах.
— Чарльз Дарвин, Происхождение человекаВ 1871 году почти не было человеческих окаменелостей доступны древние гоминины. Почти пятьдесят лет спустя предположение Дарвина подтвердилось, когда антропологи начали находить окаменелости древних гомининов с маленьким мозгом в нескольких районах Африки (список окаменелостей гоминин ). Гипотеза недавнего (в отличие от архаичного ) африканского происхождения возникла в 20 веке. «Недавнее африканское происхождение» современных людей означает «единственное происхождение» (моногенизм) и использовалось в различных контекстах как антоним полигенизма. К середине ХХ века дебаты в антропологии склонились в пользу моногенизма. Изолированные сторонники полигенизма в середине 20-го века, такие как Карлтон Кун, еще в 1962 году считали, что H. sapiens пять раз произошел от H. erectus в пяти местах.
Исторической альтернативой недавней модели происхождения является мультирегиональное происхождение современного человека, первоначально предложенное Милфордом Вулпоффом в 1980-х годах. Эта точка зрения предполагает, что происхождение анатомически современных человеческих популяций от H. erectus в начале плейстоцена 1,8 миллиона лет назад произошло в пределах постоянной популяции мира. Гипотеза неизбежно отвергает предположение о барьере бесплодия между древними евразийскими и африканскими популяциями Homo. В конце 1980-х и 1990-х годах эта гипотеза вызывала споры. Нынешняя терминология «недавнего происхождения» и «из Африки» стала актуальной в контексте этих дебатов в 1990-х годах. Первоначально рассматриваемая как противоположная альтернатива модели недавнего происхождения, мультирегиональная гипотеза в ее первоначальной «сильной» форме устарела, в то время как ее различные модифицированные более слабые варианты превратились в варианты точки зрения «недавнего происхождения» в сочетании с архаической примесью. Стрингер (2014) различает исходную или «классическую» межрегиональную модель, существовавшую с 1984 года (ее формулировка) до 2003 года, от «слабого» варианта после 2003 года, который «приблизился к модели ассимиляции».
В 1980-х годах Аллан Уилсон вместе с Ребеккой Л. Канн и Марком Стоункингом работали над генетическим датированием матрилинейный самый недавний общий предок современных человеческих популяций (получивший название «Митохондриальная Ева »). Чтобы идентифицировать информативные генетические маркеры для отслеживания истории эволюции человека, Уилсон сосредоточился на митохондриальной ДНК (мтДНК), передаваемой от матери к ребенку. Этот материал ДНК быстро мутирует, что позволяет легко отображать изменения за относительно короткое время. Открыв, что человеческая мтДНК генетически намного менее разнообразна, чем мтДНК шимпанзе, Уилсон пришел к выводу, что современные человеческие популяции недавно отделились от одной популяции, в то время как более старые человеческие виды, такие как неандертальцы и Homo erectus вымерли. С появлением археогенетики в 1990-х годах датировка митохондриальных и Y-хромосомных гаплогрупп стала возможной с некоторой уверенностью. К 1999 году оценки составляли около 150 000 лет для mt-MRCA и от 60 000 до 70 000 лет для миграции из Африки.
С 2000 по 2003 гг. Возникли разногласия по поводу митохондриальной ДНК «Mungo Man 3 » (LM3) и его возможное влияние на мультирегиональную гипотезу. Было обнаружено, что LM3 имеет большее, чем ожидалось, количество различий в последовательностях по сравнению с современной ДНК человека (CRS ). Сравнение митохондриальной ДНК с ДНК древних и современных аборигенов привело к выводу, что человек манго выходит за пределы диапазона генетических вариаций, наблюдаемых у австралийских аборигенов, и был использован для поддержки гипотезы о мультирегиональном происхождении. Повторный анализ LM3 и других древних образцов из этого района, опубликованный в 2016 году, показал, что он схож с последовательностями современных австралийских аборигенов, что несовместимо с результатами более раннего исследования.
Поскольку текущие оценки самого недавнего общего предка мужского пола («Y-хромосомный Адам » или Y-MRCA) сходятся с оценками возраста анатомически современных людей и намного предшествуют Out Гипотезы о географическом происхождении Африки по-прежнему ограничиваются Африканским континентом.
. Наибольшее количество базальных линий было обнаружено в Западе, Северо-Западе и Центральной Африке., что указывает на правдоподобность Y-MRCA, проживающего в основном регионе «Центрально-Северо-Западная Африка».
другое исследование обнаруживает вероятное размещение в «северо-западном квадранте Африканского континента» для появление гаплогруппы A1b. Отчет 2013 года о гаплогруппе A00, найденной среди людей Mbo современного западного Камеруна, также совместим с этой картиной.
Пересмотр Филогения Y-хромосомы с 2011 г. повлияла на оценки вероятного географического происхождения Y-MRCA, а также на оценки временной глубины. По той же причине будущее открытие неизвестных в настоящее время архаических гаплогрупп у живых людей снова приведет к таким пересмотрам. В частности, возможное присутствие от 1% до 4% ДНК неандертальского происхождения в евразийских геномах означает, что (маловероятное) событие обнаружения одного живого евразийского самца, демонстрирующего неандертальскую патрилинейную линию, немедленно подтолкнет вернуть T-MRCA («время до MRCA») как минимум в два раза больше текущей оценки. Однако открытие Y-хромосомы неандертальца Mendez et al. Исследование, проведенное в 2016 году, предполагает исчезновение неандертальских отцов, поскольку происхождение, выведенное из последовательности неандертальцев, выходит за пределы диапазона современных человеческих генетических вариаций. Вопросы географического происхождения станут частью дискуссии об эволюции неандертальцев от Homo erectus.
Дополнительная литература