
Войти
| Субъединица 2 комплекса распознавания источника | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | ORC2 | ||||||||
| Pfam | PF04084 | ||||||||
| InterPro | IPR007220 | ||||||||
| |||||||||
| N-конец субъединицы 3 комплекса распознавания источника (ORC) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | ORC3_N | ||||||||
| Pfam | PF07034 | ||||||||
| InterPro | IPR010748 | ||||||||
| |||||||||
| Субъединица 6 комплекса распознавания источника (ORC6) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | ORC6 | ||||||||
| Pfam | PF05460 | ||||||||
| InterPro | IPR008721 | ||||||||
| |||||||||
В молекулярной биологии комплекс распознавания происхождения (ORC ) является мульти-субъединицей ДНК связывающий комплекс (6 субъединиц), который связывается у всех эукариот и архей АТФ -зависимым образом с источниками репликации. Субъединицы этого комплекса кодируются ORC1, ORC2, ORC3, ORC4, ORC5 и ORC6 гены. ORC является центральным компонентом репликации эукариотической ДНК и остается связанным с хроматином в источниках репликации на протяжении клеточного цикла.
ORC направляет Репликация ДНК по всему геному и необходима для ее инициации. ORC, связанный в источниках репликации, служит основой для сборки пререпликационного комплекса (pre-RC), который включает Cdc6, Tah11 (он же Cdt1 ) и комплекс Mcm2 - Mcm7. Предварительная сборка RC во время G1 требуется для репликации лицензирования хромосом до синтеза ДНК во время S фазы. Регулируемое клеточным циклом фосфорилирование Orc2, Orc6, Cdc6 и MCM с помощью циклин -зависимой протеинкиназы Cdc28 регулирует инициацию ДНК репликация, включая блокирование повторной инициации в фазе G2 /M.
ORC присутствует на протяжении всего клеточного цикла, связанного с источниками репликации, но активен только в поздних митозах и ранних G1.
. В дрожжах ORC также играет роль в установлении сайленсинга в локусах mating-type Hidden MAT Left (HML) и Hidden MAT Right (HMR). ORC участвует в сборке транскрипционно молчащего хроматина на HML и HMR, рекрутируя Sir1 сайленсирующий белок в сайленсеры HML и HMR.
И Orc1, и Orc5 связывают АТФ, хотя только Orc1 обладает активностью АТФазы. Связывание АТФ с помощью Orc1 требуется для связывания ORC с ДНК и важно для жизнеспособности клеток. Активность АТФазы Orc1 участвует в формировании пре-RC. Связывание АТФ с помощью Orc5 имеет решающее значение для стабильности ORC в целом. Для привязки источника требуются только субъединицы Orc1-5 ; Orc6 необходим для поддержания сформированных pre-RC. Взаимодействия внутри ORC предполагают, что Orc2-3-6 может образовывать коровой комплекс.
 Play media Этот процесс был инициирован загрузкой Mcm2-7 на хроматиду с помощью ORC и связанных белков
Play media Этот процесс был инициирован загрузкой Mcm2-7 на хроматиду с помощью ORC и связанных белков В ORC присутствуют следующие белки:
| S. cerevisiae | S. Помбе | Д. melanogaster | Позвоночные |
|---|---|---|---|
| ORC 1-6 | ORC 1-6 | ORC 1-6 | ORC 1-6 |
| Cdc6 | Cdc18 | Cdc6 | Cdc6 |
| Cdt1 / Tah11 / Sid2 | Cdt1 | DUP | Cdt1 / RLF-B |
| Mcm2 | Mcm2 / Cdc19 / Nda1 | Mcm2 | Mcm2 |
| Mcm3 | Mcm3 | Mcm3 | Mcm3 |
| Cdc54 / Mcm4 | Cdc21 | DPA | Mcm4 |
| Cdc46 / Mcm5 | Mcm5 / Nda4 | Mcm5 | Mcm5 |
| Mcm6 | Mcm6 / Mis5 | Mcm6 | Mcm6 |
| Cdc47 / Mcm7 | Mcm7 | Mcm7 | mcm7 |
Археи имеют упрощенную версию ORC, Mcm и, как следствие, комбинированный pre-RC. Вместо использования шести разных белков mcm для формирования псевдосимметричного гетерогексамера, все шесть субъединиц в MCM архей одинаковы. У них обычно есть несколько белков, гомологичных как Cdc6, так и Orc1, некоторые из которых выполняют функцию обоих. В отличие от эукариотических орков, они не всегда образуют комплекс. Фактически, они имеют расходящиеся сложные структуры, когда они формируются. Sulfolobus islandicus также использует гомолог Cdt1 для распознавания одного из источников его репликации.
Автономно реплицирующиеся последовательности (ARS), впервые обнаруженные в почкующиеся дрожжи являются неотъемлемой частью успеха ORC. Эти последовательности 100-200 п.н. способствуют репликационной активности во время S-фазы. ARS могут быть размещены в любом новом месте хромосом почкующихся дрожжей и будут способствовать репликации из этих сайтов. Считается, что высококонсервативная последовательность из 11 пар оснований (известная как элемент A) важна для функции происхождения у почкующихся дрожжей. ORC был первоначально идентифицирован по его способности связываться с элементом A ARS у почкующихся дрожжей.
Клетки животных содержат гораздо более загадочную версию ARS, при этом пока не обнаружено консервативных последовательностей. Однако в клетках животных точки начала репликации собираются в пучки, называемые кластерами репликона. Репликоны каждого кластера имеют одинаковую длину, но отдельные кластеры имеют репликоны разной длины. Все эти репликоны имеют сходные основные остатки, с которыми связывается ORC, которые во многом имитируют консервативный элемент A из 11 пар оснований. Все эти кластеры одновременно активируются во время фазы S.
ORC необходим для загрузки комплексов MCM (Pre-RC ) на ДНК. Этот процесс зависит от ORC, Cdc6 и Cdt1, включая несколько событий набора, контролируемых ATP. Во-первых, ORC и Cdc6 образуют комплекс по происхождению ДНК (отмечен областями типа ARS). Новые комплексы ORC / Cdc6 затем привлекают молекулы Cdt1 / Mcm2-7 к сайту. Как только этот массивный комплекс ORC / Cdc6 / Cdt1 / Mcm2-7 (OCCM) образуется, молекулы ORC / Cdc6 / Cdt1 работают вместе, загружая Mcm2-7 на саму ДНК посредством гидролиза АТФ по Cdc6. Фосфорилирующая активность Cdc6 зависит как от ORC, так и от исходной ДНК. Это приводит к тому, что Cdt1 имеет пониженную стабильность на ДНК и выпадает из комплекса, что приводит к нагрузке Mcm2-7 на ДНК. Структура ORC, MCM, а также промежуточного комплекса OCCM была определена.
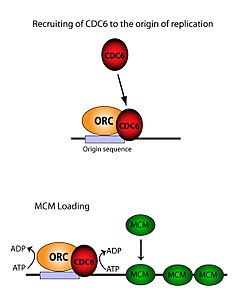

Хотя ORC состоит из шести дискретных субъединиц, только одна из них оказалась значительный - ORC1. Исследования in vivo показали, что Lys -263 и Arg -367 являются основными остатками, ответственными за точную загрузку ORC. Эти молекулы представляют собой упомянутый выше ARS. ORC1 взаимодействует с ATP и этими основными остатками, чтобы связать ORC с исходной ДНК. Было установлено, что это происходит задолго до репликации и что сам ORC уже связан с исходной ДНК к моменту любой загрузки Mcm2-7. При первой загрузке Mcm2-7 он полностью окружает ДНК, и активность геликазы ингибируется. В S-фазе комплекс Mcm2-7 взаимодействует с кофакторами геликазы Cdc45 и GINS для выделения одного Нить ДНК, разверните ориджин и начните репликацию по хромосоме. Чтобы иметь двунаправленную репликацию, этот процесс происходит дважды в источнике. Оба события нагрузки опосредуются одним ORC посредством процесса, идентичного первому.
Всесторонний обзор репликации молекулярной ДНК