
Войти
 Эукариотическая Репликация ДНК
Эукариотическая Репликация ДНК Репликация эукариотической ДНК - это консервативный механизм, который ограничивает Репликация ДНК до одного раза за клеточный цикл. Репликация эукариотической ДНК хромосомной ДНК является центральной для дублирования клетки и необходима для поддержания репликации ДНК эукариот генома.
представляет собой действие ДНК-полимераз, синтезирующие цепи ДНК, комплементарную исходную цепь матрицы. Для синтеза ДНК двухцепочечная ДНК разматывается ДНК геликазами перед полимеразами, образуя репликационную вилку, содержащую две одноцепочечные матрицы. Процессы репликации позволяют копировать одиночную двойную спираль ДНК в две спирали ДНК, которые разделяются на дочерние клетки при митозе. Основные ферментативные функции, выполняемые на репликационной вилке, хорошо сохраняются от прокариот до эукариот, но репликационный аппарат в репликации эукариотической ДНК представляет собой гораздо более крупный комплекс, координирующий многие белки на этом участке. репликации, образуя реплисому.
Реплисома отвечает за копирование всей геномной ДНК в каждой пролиферативной клетке. Этот процесс позволяет передать информацию о родительской клетке к дочерней нейке, необходимой для всех организмов. Большая часть клеточного цикла построена вокругобеспечения того, чтобы репликация ДНК происходила без ошибок.
В G1фазе клеточного цикла запускаются процессы регуляции репликации ДНК.. У эукариот подавляющая часть синтез ДНК происходит во время S фазы клеточного цикла, и весь геном должен раскручен и продублирован для образования двух дочерних копий. В течение G2 любые поврежденные ДНК или ошибки репликации исправляются. Наконец, одна копия генома отделяется от каждой дочерней клетки митозе или М-фазе. Каждая изэтих дочерних копий содержит одну родительскую цепь дуплексной ДНК и одну родительскую антипараллельную цепь.
Этот механизм сохраняется от прокариот до эукариот и известен как полуконсервативная репликация ДНК. Процесс полуконсервативной репликации для сайта репликации ДНК представляет вилкообразную ДНК, вилку репликации, где спираль ДНК открыта или раскручена, обнажая неспаренные ДНК нуклеотиды для распознавания и спаривания оснований включения нуклеотидов вдвухцепочечную ДНК.
Инициирование репликации эукариотической ДНК - это первая стадия синтеза ДНК, на которой двойная спираль ДНК разматывается и начальное событие праймирования ДНК-полимеразой происходит на ведущей цепи. Событие прайминга на отстающую нити устанавливает вилку репликации. Праймирование спирали ДНК заключается в синтезе праймера РНК, позволяющем синтезировать ДНК-полимеразой α.Праймирование происходит один раз в начале ведущей нити и в начале каждого фрагмента Окадзаки на отстающую нити.
Репликация ДНК действует через последовательность, называемых точками репликации, и эукариотические клетки множественные точки начала репликации. Чтобы инициировать репликацию ДНК, несколько репликативных белков собираются и отделяются от этих репликативных источников. Отдельные факторы, описанные ниже, работают вместе, чтобы управлять образованием пререпликационного комплекса (до RC), ключевого промежуточного звена в процессе инициации репликации.
Ассоциация комплекса распознавания ориджина (ORC) с ориджином репликации привлекает белок цикла 6 деления клетки (Cdc6), чтобы сформировать платформу для загрузки комплексных поддерживающих минихромосом ( Mcm 2-7) белков, чему способствует лицензирование хроматина и белок-фактор репликации 1 ДНК (Cdt1). ORC, Cdc6 и Cdt1 вместе необходимы для стабильнойассоциации комплекса Mcm2-7 с репликативными источниками во время фазы G 1 клеточного цикла.
Эукариотические источники репликации контролируют образование ряда белковых комплексов, которые приводят к сборке двух двунаправленных вилок репликации ДНК. Эти события запускают образование пререпликационного комплекса (пре-RC) в источнике репликации. Этот процесс происходит на стадии G 1 клеточного цикла. Формирование пре-RCвключает упорядоченную сборку многих факторов репликации, включая комплекс распознавания ориджина (ORC), белок Cdc6, белок Cdt1 и поддерживающие белки минихромосомы (Mcm2-7). Как только пре-RC сформирован, активация комплекса запускается двумя киназами, циклин-зависимой киназой 2 (CDK) и Dbf4-зависимой киназой (DDK), которые обеспечивают перевести пре-RC в комплекс инициации до инициации репликации ДНК. Этот переход включает упорядоченную сборку дополнительных фактороврепликации для раскручивания ДНК и накопления множественных полимераз эукариотической ДНК вокруг раскрученной ДНК. Центральным в вопросе о том, как двусторонние репликационные вилки устанавливаются в источнике репликации, механизм, с помощью которого ORC рекрутирует два встречных комплекса Mcm2-7 в каждую точку начала репликации, чтобы сформировать пререпликационный комплекс.
Первым шагом в сборке пререпликационного комплекса (pre-RC):связывание комплекса распознавания ориджина (ORC) с ориджином репликации. В позднем митозе белок Cdc6 присоединяется к связанному ORC с последующим связыванием комплекса Cdt1-Mcm2-7. ORC, Cdc6 и Cdt1 все необходимы для загрузки комплекса из шести белков минихромосомы (Mcm 2-7) на ДНК. ORC представляет собой шести-субъединичный белковый комплекс Orc1p-6, который выбирает репликативные сайты инициации ДНК для инициации репликации, и связывание ORC с хроматином регулируется в рамкахклеточного цикла. Как правило, функция и размер субъединиц ORC сохраняются во многих геномах эукариот, с той разницей, что их сайты связывания ДНК расходятся.
Наиболее широко изученным комплексом распознавания ориджина является комплекс Saccharomyces cerevisiae или дрожжей, который, как известно, связывается с автономно реплицирующейся последовательностью (ARS). ORC S. cerevisiae специфически взаимодействует как с элементами A, так и с B1 дрожжевыми источникамирепликации, охватывая область из 30 пар основ. Для связывания последовательностей требуется АТФ.
. Определена атомная структура ORC S. cerevisiae, связанного с ДНК ARS. Orc1, Orc2, Orc3, Orc4 и Orc5 окружают элементы посредством двух типов специфичных оснований, изгибают ДНК в элементе A. Все пять субъединиц контактируют с сахарно-фосфатным остальным в точных точках элемента A, образуя плотный захват без специфичности основания. Orc1 и Orc2 контактируют с малой бороздкой элементаA, в то время как домен крылатой спирали Orc4 контактирует с метильными группами инварианта Ts в большой бороздке элемента A через спираль вставки (IH). Отсутствие этого IH у многоклеточных животных отчасти объясняет отсутствие специфической последовательности в ORC человека. ДНК ARS также изгибается в элементе B1 за счет взаимодействия с Orc2, Orc5 и Orc6. Изгибание исходной ДНК с помощью ORC, по-видимому, эволюционно консервативно, что позволяет предположить, что это может быть механизмвключения комплекса Mcm2-7.
Когда ORC связывается с ДНК в начале репликации, он служит каркасом для сбора других факторов инициации пререпликативного комплекса. Сборка этого пререпликативного комплекса во время стадии G 1 клеточного цикла требуется до активации репликации ДНК во время S-фазы. Удаление по крайней мере части комплекса (Orc1) из хромосомы в метафазе является частью регуляции ORC млекопитающих, чтобы уничтожить образование пререпликативного комплекса дозавершения метафазы, устранено. 73>
Связывание белка цикла 6 цикла деления клеток (Cdc6) с комплексом распознавания ориджина (ORC) является важным этапом в сборке пререпликации комплекса (пре-RC) в истоках репликации. Cdc6 связывается с ORC на ДНК АТФ-зависимым образом, что вызывает изменение характера связывания источника, для которого требуется Orc1 АТФаза. Cdc6 требует ORC для связывания с хроматином и, в свою очередь, необходим для связываниягептамера Cdt1-Mcm2-7 с хроматином. Комплекс ORC-Cdc6 образует кольцевую структуру и подобен другим АТФ-зависимым белковым машинам. Уровни и активность Cdc6 регулирует частоту, используя источники репликации в течение клеточного цикла.
Белок лицензирования хроматина и фактора репликации ДНК 1 (Cdt1) требуется для лицензирования хроматина для репликации ДНК. У S. cerevisiae Cdt1 облегчает загрузку комплекса Mcm2-7 по одному на хромосому за счет стабилизациилевой структуры с открытым кольцом одиночного гексамера Mcm2-7. Было показано, что Cdt1 ассоциируется с С-концом Cdc6, чтобы кооперативно ассоциации белков Mcm с хроматином. Крио-ЭМ структура комплекса OCCM (ORC-Cdc6-Cdt1-MCM) показывает, что Cdt1-CTD взаимодействует с Mcm6-WHD. У многоклеточных животных активность Cdt1 во время клеточного цикла жестко регулируется за счет его ассоциации с белком геминином, который ингибирует активность Cdt1 во время S-фазы, чтобы предотвратитьповторную репликацию ДНК и предотвращает ее убиквитинирование и последующий протеолиз.
Белки поддержания минихромосомы (Mcm) были названы в честь генетического скрининга мутантов инициации репликации ДНК в S. cerevisiae, которые обеспечивают стабильность плазмиды в специфическом для ARS методе. Mcm2, Mcm3, Mcm4, Mcm5, Mcm6 и Mcm7 образуют гексамерный комплекс, который имеет структуры открытого кольца с зазором между Mcm2 и Mcm5.Сборка белков Mcm на хроматин требует скоординированной функции распознавания комплекса источника (ORC), Cdc6 и Cdt1. После того, как белки Mcm были загружены в хроматин, ORC и Cdc6 могут быть удалены из хроматина без предотвращения последующей репликации ДНК. Это наблюдение предполагает, что первичная роль пререпликационного комплекса является правильной загрузкой белков Mcm.
 Двойной гексамер Mcm2-7 расположен в ориентации «голова к голове» (NTD-to-NTD). Каждое гексамерное кольцо слегканаклонено, скручено и смещено относительно друг друга. Верхняя панель, виды сбоку. Нижняя панель, CTD-изображение.
Двойной гексамер Mcm2-7 расположен в ориентации «голова к голове» (NTD-to-NTD). Каждое гексамерное кольцо слегканаклонено, скручено и смещено относительно друг друга. Верхняя панель, виды сбоку. Нижняя панель, CTD-изображение. Белки Mcm на хроматине образуют двойной гексамер с двумя кольцами, слегка наклоненными, скрученными и смещенными от центра, чтобы создать перегиб в центральном канале, где связанная ДНК захватывается интерфейс двух колец. Каждое гексамерное кольцо Mcm2-7 служит каркасом для сборки реплисомы, а затем ядром каталитической геликазы CMG (Cdc45-MCM-GINS), который является основнымкомпонентом реплисомы. Каждый белок Mcm отличается от других. У всех эукариот есть ровно шесть белков Mcm, каждый из которых попадает в один из новых классов (Mcm2-7), что указывает на то, что каждый белок Mcm выполняет уникальную функцию функции.
Поддерживающие белки минихромосомы необходимы для активности ДНК-геликазы. Инактивация любого из шести белков Mcm во время S-фазы предотвращает дальнейшее развитие репликационной вилки, предполагаемая, что геликаза может рециклироваться идолжна собираться в точках репликации. Наряду с хеликазной активностью комплекса поддерживающего белка минихромосомы, этот комплекс также обладает свойствами с ним АТФазной активностью. Мутация в любом из шести белков Mcm снижает консервативные сайты связывания АТФ, что указывает на то, что гидролиз АТФ является скоординированным фактом, в котором участвуют все шесть субъединиц комплекса Mcm. Исследования показали, что внутри белкового комплекса Mcm есть большие каталитические пары белков Mcm,которые вместе для гидролиза АТФ. Например, Mcm3, но не Mcm6 может активировать активность Mcm6. Эти исследования, подтвержденные крио-ЭМ-структурами комплексов Mcm2-7, предполагают, что комплекс Mcm представляет собой гексамер с Mcm3 рядом с Mcm7, Mcm2 следующим в Mcm6 и Mcm4 рядом с Mcm5. Обе члены каталитической пары вносят свой вклад в конформацию, которая обеспечивает связывание и гидролиз АТФ, а смесь активных и неактивныхсубъединиц создает скоординированную активность АТФазы, которая позволяет белковому комплексу Mcm завершить связывание и гидролиз АТФ в.
ядерная локализация поддерживающих белков минихромосомы регулируется в почкующихся дрожжевых клетках. Белки Mcm присутствуют в ядре на стадии G 1 и S-фазе клеточного цикла, но экспортируются в цитоплазму во время G 2 ступень и фаза М. Полный и интактный комплекс Mcm из шести субъединиц необходим для проникновенияв ядро клетки. У S. cerevisiae ядерному экспорту активность циклин-зависимой киназы (CDK). Белки Mcm, которые связаны с хроматином, защищены от механизма экспорта CDK из-за отсутствия доступа к CDK.
На стадии G 1 клетки В цикле факторы инициации репликации, комплекс распознавания ориджина (ORC), Cdc6, Cdt1 и белковый комплекс поддержания минихромосомы (Mcm) последовательно связываются с ДНК с образованием комплекса пререпликации (pre-RC).При переходе от стадии G 1 к S-фазе клеточного цикла, S-фаза-специфичная циклин-зависимая протеин киназа (CDK) и Cdc7 / Dbf4 киназа ( DDK) преобразует pre-RC в активную вилку репликации. Во время этого преобразования пре-RC разбирается с потерей Cdc6, создавая комплекс инициации. Помимо связывания белков Mcm, белок цикла деления клеток 45 (Cdc45) также важен для инициации репликации ДНК. Исследования, что Mcm является критическим для загрузки Cdc45 на хроматин, иэтот комплекс образует Mcm, так и Cdc45, образует в начале S-фазы клеточного цикла. Cdc45 нацелен на белковый комплекс Mcm, который загружен на хроматин, в качестве компонента пре-RC в начале репликации во время стадии G 1 клеточного цикла.
Белок цикла деления клетки 45 (Cdc45) является критическим компонентом для превращения пререпликативного комплекса в комплекс инициации. Белок Cdc45 собирается в ориджинах репликации для инициации и необходимости началарепликации в Saccharomyces cerevisiae, и играет важную роль во время элонгации. Таким образом, Cdc45 играет центральную роль как в фазах инициации, так и в фазах элонгации репликации хромосомной ДНК.
Cdc45 ассоциируется с хроматином после начала инициации на поздней стадии G 1 и во время фазы S хромосомной ДНК. клеточный цикл. Cdc45 физически ассоциируется с Mcm5 и демонстрирует генетические различия с пятью членами семейства генов Mcm и геном ORC2. Загрузка Cdc45 нахроматин имеет решающее значение для загрузки других различных белков репликации, включая ДНК-полимеразу α, ДНК-полимеразу, белок репликации A (RPA) и ядерный антиген пролиферирующих клеток. (PCNA) на хроматин. В системе без ядра Xenopus было установлено, что Cdc45 необходим для раскручивания плазмидной ДНК. Безъядерная система Xenopus также демонстрирует, что раскручивание ДНК и прочное связывание RPA с хроматином происходит только в присутствии Cdc45.
Связывание Cdc45 с хроматином зависит от активности киназы Clb-Cdc28, а также функциональных Cdc6 и Mcm2, что предполагает, что Cdc45 связывается с пре-RC после активации S-фазы циклин-зависимых киназ (CDK). Как показывает время и зависимость CDK, связывание Cdc45 с хроматином является критическим для инициации репликации ДНК. Во время фазы S Cdc45 физически взаимодействует с белками Mcm на хроматине; однако диссоциация Cdc45 от хроматина происходит медленнее, чем диссоциация Mcm, что указываетна то, что белки высвобождаются по разным механизмам.
Шесть поддерживающих белков минихромосом и Cdc45 важны во время инициации и удлинение для движения вилок репликации и раскручивания ДНК. GINS важны для взаимодействия Mcm и Cdc45 в источнике репликации во время инициации, а затем в ответвлениях репликации по мере развития реплисомы. Комплекс GINS состоит из четырех небольших белков Sld5 (Cdc105), Psf1 (Cdc101), Psf2 (Cdc102) и Psf3 (Cdc103), GINS представляет собой«go, ichi, ni, san», что означает «5, 1, 2, 3.. ». на японском языке. Cdc45, Mcm2-7 и GINS вместе образуют геликазу CMG, репликативную геликазу реплисомы. Хотя комплекс Mcm2-7 сам по себе имеет слабую геликазную активность, Cdc45 и GINS необходимы для устойчивой геликазной активности
Mcm10 важен для репликации хромосомы и имеет значение с минихромосомной геликазой 2-7, которая поддерживает загружается в неактивной форме в начале репликации ДНК. Mcm10 также шапероны каталитической ДНК-полимеразы α и помогает стабилизировать полимеразу на репликационных вилках.
В начале S-фазы пререпликативная Комплекс должен быть активирован двумя киназами, специфичными для S-фазы, чтобы образовать комплекс инициации в ориджине репликации. Одна киназа представляет собой киназу Cdc7-Dbf4, называемую Dbf4-зависимой киназой (DDK), и другую - циклин-зависимой киназой (CDK). Анализ связывания хроматина Cdc45 у дрожжей и Xenopusпоказал, что последующим событием действия CDK является загрузка Cdc45 на хроматин. Cdc6, как предполагалось, является мишенью CDK, из-за ассоциации между Cdc6 и CDK и CDK-зависимого фосфорилирования Cdc6. Считается, что CDK-зависимое фосфорилирование Cdc6 необходимо для перехода в S-фазу.
Как каталитические субъединицы DDK, Cdc7, так и активаторный белок, Dbf4, консервативны у эукариот и необходимы для наступления S фазы клеточного цикла. И DDK, и Cdc7необходимы для загрузки Cdc45 в точки начала репликации хроматина. Мишенью для связывания киназы DDK является комплекс Mcm, возможно, Mcm2. DDK нацелен на комплекс Mcm, и его фосфорилирование приводит к возможной активации активности геликазы Mcm.
Sld3, Sld2 и Dpb11 взаимодействуют со многими белками репликации. Sld3 и Cdc45 образуют комплекс, который ассоциирован с до-RC в ранних источниках репликации даже в фазе G1 1 и с более поздниминовыми репликациями в S-фазе взаимно Mcm-зависимым образом. Dpb11 и Sld2 взаимодействуют с полимеразой, и эксперименты по перекрестному связыванию показали, что Dpb11 и полимераза совместно преципитируют в S-фазе и связываются с источниками репликации.
Sld3 и Sld2 фосфорилируются CDK, что позволяет двум репликативным белкам привязать к Dpb11. Dpb11 имел две пары доменов С-конца BRCA1 (BRCT), которые известны как фосфопептид-связывающие домены. N-концевая пара доменов BRCT связывается сфосфорилированным Sld3, а C-концевая пара связывается с фосфорилированным Sld2. Оба этих взаимодействия необходимы для CDK-зависимой активации почкования ДНК у дрожжей.
Dpb11 также взаимодействует с GINS и участвует в этапах инициации и удлинения репликации хромосомной ДНК. GINS является одним из белков репликации, обнаруженных в ответвлениях репликации, и образует комплекс с Cdc45 и Mcm.
Эти зависимые от фосфорилирования взаимодействия между Dpb11, Sld2 и Sld3 необходимы дляCDK-зависимой активации репликации ДНК, и при использовании сшивающих реагентов в некоторых экспериментах был идентифицирован хрупкий комплекс, называемый предварительной загрузкой сложный (до LC). Этот комплекс содержит Pol ɛ, GINS, Sld2 и Dpb11. Обнаружено, что пре-LC формирует передовую ассоциацию с ориджинами CDK-зависимым и DDK-зависимым образом, и активность CDK регулирует инициацию репликации ДНК посредством образования пре-LC.
 Комплекс эукариот реплисома иассоциированные белки. В отстающей цепи возникает петля
Комплекс эукариот реплисома иассоциированные белки. В отстающей цепи возникает петля . Образование пререпликативного комплекса (пре-RC) потенциальные сайты для инициации репликации ДНК. В соответствии с комплексом поддержания минихромосомы, окружающая двухцепочечная ДНК, образование пре-RC не приводит к немедленному раскручиванию исходной ДНК или привлечению ДНК-полимераз. Вместо этого пре-RC, который образуется во время G 1 клеточного цикла, активируется только для раскручивания ДНК и инициации репликациипосле того, как клетки переходят из G 1 в S фаза клеточного цикла.
Как только инициирующий комплекс образуется и клетки переходят в S-фазу, комплекс становится реплисомой. Комплекс реплисом эукариот отвечает за координацию репликации ДНК. Репликация на ведущей и отстающей цепях осуществляется ДНК-полимеразой ε и ДНК-полимеразой δ. Многие факторы реплисомы, включая Claspin, And1, загрузчик зажима фактора репликации C и комплекс вилки, ответственны за регуляцию функций полимеразыи координации синтеза ДНК с раскручиванием цепи, матрицы комплексом Cdc45-Mcm-GINS. По мере разматывания ДНК число скручивания уменьшается. Чтобы компенсировать это, число корч увеличить, вводя в ДНК положительные суперспирали. Эти суперспирали остановили бы репликацию ДНК, если бы они не были удалены. Топоизомеразы ответственны за удаление этих суперспиралей перед вилкой репликации.
Реплисома отвечает за копирование всей геномной ДНК в каждой пролиферативнойклетке. Реакции спаривания оснований и образования цепей, образующие дочернюю спираль, катализируются ДНК-полимеразами. Эти ферменты перемещаются вдоль одноцепочечной ДНК и позволяют удлинить зарождающуюся цепь ДНК, «считывая» цепочку-матрицу и включение правильных пуриновых азотистых оснований, аденин и гуанин, и пиримидин азотистые основания, тимин и цитозин. Активированные свободные дезоксирибонуклеотиды существуют в клетке в видедезоксирибонуклеотидтрифосфатов (dNTP). Эти свободные нуклеотиды добавляются к открытой 3'-гидроксильной группе на последнем включенном нуклеотиде. В этой реакции из свободного dNTP высвобождается, генерируется пи энергия для реакции полимеризации и обнажая 5'-монофосфат, который ковалентно связывается с 3'-кислородом. Кроме того, неправильно вставленные нуклеотиды могут быть удалены и заменены правильными нуклеотидами в энергетически выгодной реакции. Это свойство имеет жизненно важноезначениедля правильной проверки и исправления ошибок, содержащих во время репликации ДНК.
Репликационная вилка - это соединение между недавно разделенными цепями матрицы, известными как ведущая и отстающая цепи, и двухцепочечной ДНК. ДНК-полимеразы синтезируют ДНК в направлении от 5 'к 3' относительно вновь синтезированной цепи, и все ДНК-полимеразы синтезируют ДНК в противоположных направлениях между двумя новыми цепями в репликационной вилке. Дальнейшаякоординация требуется во время репликации ДНК. Две репликативные полимеразы синтезируют ДНК в противоположных ориентациях. Полимераза непрерывно синтезирует ДНК на «ведущей» цепи ДНК, поскольку она указывает в том же направлении, что и раскручивание ДНК реплисомой. Напротив, полимераза синтезирует ДНК на «отстающей» цепи, которая является противоположной цепью матрицы ДНК, фрагментированным или прерывистым образом.
Прерывистые участки продуктов репликации ДНК на отстающей цепи известныкак фрагменты Окадзаки и составляют от 100 до 200 оснований в длину на эукариотических вилках репликации. Отстающая цепь обычно содержит более длинные участки одноцепочечной ДНК, покрытой одноцепочечными связывающими белками, которые позволяют предотвратить образование вторичной структуры. У эукариот эти одноцепочечные связывающие белки представляет собой гетеротримерный комплекс, известный как репликационный белок A (RPA).
Каждому фрагменту Окадзаки предшествует праймер РНК,который вытесняется цепочкой следующего фрагмента Окадзаки в процессе синтеза. РНКаза H распознает гибриды ДНК: РНК, созданная с использованием праймеров РНК, отвечает за их удаление из реплицированной цепи, оставляя после себя соединение праймер: матрица. ДНК-полимераза определяет эти сайты и удлиняет разрывы, оставленные удалением праймера. В эукариотических клетках небольшое количество ДНК непосредственно перед праймером РНК также смещается, создавая лоскутную структуру. Затем этотлоскут расщепляется эндонуклеазами. В репликационной вилке разрыва ДНК после удаления лоскута закрывается ДНК-лигазой I, которая восстанавливает разрывы, оставшиеся между 3'-ОН и 5'фосфатом вновь синтезированной цепи. Из-за относительно короткой природы эукариотического фрагмента Окадзаки синтез репликации ДНК, происходящий с перерывами на отстающей цепи, менее эффективен и требует больше времени, чем синтез ведущей цепи. Синтез ДНК завершается после удаления всех праймеров РНК и заделкизазубрин.
 Изображение репликации ДНК на вилке репликации. a : нити шаблона, b : ведущая нить, c : отстающая нить, d : репликационная вилка, e : Праймер РНК, f : фрагмент Окадзаки
Изображение репликации ДНК на вилке репликации. a : нити шаблона, b : ведущая нить, c : отстающая нить, d : репликационная вилка, e : Праймер РНК, f : фрагмент Окадзаки Во время репликации ДНК реплисома раскручивает родительскую дуплексную ДНК в двухцепочечную репликационную вилку матрицы ДНК в Направление от 5 до футов 3 футов. Ведущая нить - это нить-матрица, которая реплицируется в том женаправлении, что и движение репликационной вилки. Это позволяет синтезировать вновь синтезированную цепь, комплементарную исходную цепь, от 5 'к 3' в том же направлении, что и движение репликационной вилки.
После того, как праймер РНК был добавлен примазой к 3 На конце ведущей цепи синтеза ДНК будет продолжаться в направлении от 3 'до 5' по отношению к ведущей цепи без прерывания. ДНК-полимераза будет непрерывно добавлять нуклеотиды к матричной цепи, поэтому для синтеза ведущей цепитребуется один праймер и она обладает непрерывной активностью ДНК-полимеразы.
Репликация ДНК на отстающая цепь прерывистый. При синтезе отстающей цепи движение ДНК-полимеразы в противоположном репликационной вилке, требует использования нескольких праймеров РНК. ДНК-полимераза будет синтезировать короткие фрагменты ДНК, называемые фрагментами Окадзаки, которые добавляются к 3'-концу праймера. Эти фрагменты могут иметь длину от100 до 400 нуклеотидов у эукариот.
В конце синтез фрагмента Окадзаки ДНК-полимераза проходит в предыдущий фрагмент Окадзаки и смещает его 5'-конец, обеспечива праймер РНК и небольшой сегмент ДНК. Это генерирует однонитевой лоскут РНК-короткие ДНК, который необходимо расщепить, разрыв между двумя фрагментами Окадзаки должен быть запломбирован ДНК-лигазой I. Этот процесс известен как созревание фрагмента Окадзаки и может осуществляться двумя способами: один механизм процессов закрылки, адругой касается длинных закрылков. ДНК-полимераза δ способна заменить от 2 до 3 нуклеотидов ДНК или РНК перед ее полимеризацией, создавая короткий субстрат «лоскута» для Fen1, который может удалять нуклеотиды из лоскута, по одному нуклеотиду на время.
Повторяя циклы этого процесса, ДНК-полимераза δ и Fen1 могут координировать удаление праймеров РНК и оставлять разрыв ДНК на отстающую цепи. Это предположительно, что он строго регулируется и не вызывает больших лоскутов, которыенеобходимы иссекать. В случае нарушения регуляции активности Fen1 / ДНК-полимеразы δ-клетка использует альтернативный механизм для создания и обработки длинных створок с помощью Dna2, который обладает как геликазной, так и нуклеазной активностями. Нуклеазная активность Dna2 необходима для удаления этих длинных лоскутов, оставляя более короткий лоскут для обработки Fen1. Электронная микроскопия показывают, что нагрузка нуклеосом на отстающую цепь происходит очень близко к месту синтеза.Таким образом, созревание фрагмента Окадзаки представляет собой эффективный процесс, который происходит сразу после зарождающейся ДНК.
После того, как репликативная геликаза раскрутила дуплекс родительской ДНК, открыли две одноцепочечные ДНК-матрицы, репликативные полимеразы необходимы для генерации двух копий родительского генома. Функция ДНК-полимеразы является узкоспециализированной и выполняет репликацию определенных шаблонов и в узкихлокализациях. На вилке репликации эукариот есть три различных репликативных полимеразных комплекса, которые вносят вклад в репликацию ДНК: полимераза α, полимераза δ и полимераза ε. Эти три полимеразы необходимы для жизнеспособности клетки.
ДНК-полимеразам требуется праймер для начала синтеза ДНК, полимераза α (Pol α) как репликативная примаза. Pol α связан с РНК-примазой, этот комплекс выполняет задачу синтеза праймера, который содержит короткий 10-нуклеотидный участок РНК, за которымследуют от 10 до 20 основ ДНК. Важно, что это прайминговое действие происходит при инициации репликации в ориджинах, чтобы начать синтез ведущей цепи, а также на 5'-конце каждого фрагмента Окадзаки на отстающей цепи.
Однако Pol α не может продолжать репликацию ДНК и должен быть заменен другой полимеразой для продолжения синтеза ДНК. Переключение полимеразы требует загрузчиков зажимов, и было доказано, что нормальная репликация ДНК требует скоординированных действий всех трехДНК-полимераз: Pol α для праймингового синтеза, Pol ε для репликации ведущей цепи и Pol δ, который постоянно загружается, для генерации Фрагменты Окадзаки во время синтеза отстающей цепи.
ДНК геликазы и полимеразы должны оставаться в тесном контакте на вилке репликации. Если раскрутка происходит слишком далеко до синтеза, обнажаются большие участки одноцепочечной ДНК. Это может активировать сигнализацию повреждения ДНК или вызвать процессырепарации ДНК. Чтобы предотвратить эти проблемы, Реплисома эукариот содержит специализированные белки, предназначенные для регулирования активности геликазы перед репликационной вилкой. Эти белки также обеспечивают стыковочные сайты для физического взаимодействия между геликазами и полимеразами, тем самым гарантируя, что раскручивание дуплекса связано с синтезом ДНК.
Для функционирования ДНК-полимераз необходимо размотать двухцепочечную спираль ДНК, чтобы обнажить две одноцепочечныеДНК-шаблоны для репликации. ДНК-геликазы ответственны за раскручивание геномной ДНК во время фазы S, что свидетельствует об их локализации в репликационной вилке.
Загрузка белков Mcm может происходить только во время G 1 клеточный цикл, и нагруженный комплекс затем активируется во время фазы S путем рекрутирования белка Cdc45 и комплекса GINS с образованием активной геликазы Cdc45-Mcm-GINS (CMG) в ответвлениях репликации ДНК. Активность МКМ требуется на протяжении фазы S для репликации ДНК. Множество регуляторных факторов собираются вокруг геликазы CMG, образуя «комплекс реплисомной прогрессии», который связывается с ДНК-полимеразами с образованием реплисомы эукариот, структура которой все еще довольно плохо определена по сравнению с ее бактериальным аналогом.
Изолированная геликаза CMG и комплекс реплисомной прогрессии содержат комплекс с одним кольцом белка Mcm, что позволяет предположить, что загруженный двойной гексамер белков Mcm в начале может быть разорваны на два одиночных гексамерных кольца как часть процесса инициации, причем каждое кольцо комплекса белка Mcm формирует ядро геликазы CMG в двух репликационных вилках, установленных от каждого ориджина. Полный комплекс CMG необходим для раскручивания ДНК, а комплекс CDC45-Mcm-GINS является функциональной ДНК-геликазой в эукариотических клетках.
Комплекс CMG действует с реплисомой посредством взаимодействия с белками Ctf4 и And1. Белки Ctf4 / And1 взаимодействуют как с комплексом CMG, так и с ДНК-полимеразой α. Ctf4 представляет собойдополнительный фактор полимеразы α, который необходим для привлечения полимеразы α к источникам репликации.
белки Mrc1 / Claspin соединяют синтез ведущей цепи с комплексом CMG геликазная активность. Mrc1 взаимодействует с полимеразой ε, а также с белками Mcm. Важность этой прямой связи между геликазой и полимеразой ведущей цепи подчеркивается результатами вкультивируемых клетках человека, где Mrc1 / Claspin необходим для эффективного прогрессированиярепликационной вилки. Эти результаты предполагают, что для эффективной репликации ДНК также требуется соединение геликаз и синтез ведущей цепи...
ДНК-полимеразам требуются дополнительные факторы для поддержки репликации ДНК. ДНК-полимеразы имеют полузамкнутую «ручную» структуру, которая позволяет полимеразе загружаться в ДНК и начинатьперемещаться. Эта структура позволяет ДНК-полимеразе удерживать матрицу одноцепочечной ДНК, включатьдНТФ в активный сайт и высвобождать вновь образованную двухцепочечную ДНК. Однако структура ДНК-полимераз не позволяет осуществлять непрерывное стабильное взаимодействие с матричной ДНК.
Для усиления взаимодействия между полимеразой и матричной ДНК, скользящие зажимы ДНК связываются с полимеразой, чтобы способствовать процессивность репликативной полимеразы. У эукариотскользящий зажим представляет собой кольцевую структуру гомотримеров, известную как ядерный антигенпролиферирующих клеток (PCNA). Кольцо PCNA имеет полярность с поверхностями, которые взаимодействуют с ДНК-полимеразами и надежно прикрепляют их к матрице ДНК. PCNA-зависимая стабилизация ДНК-полимераз оказывает существенное влияние на репликацию ДНК, поскольку PCNA могут увеличивать процессивность полимеразы до 1000 раз. PCNA является важным кофактором и отличается тем, что являетсяодной из наиболее распространенных платформ взаимодействия в реплисоме для приспособления множества процессовна вилке репликации, поэтому PCNA также рассматривается как регуляторный кофактор для ДНК-полимераз.
PCNA полностью окружает цепочку матрицы ДНК и должен быть загружен в ДНК на вилке репликации. Загрузка PCNA в ведущую цепь происходит нечасто, поскольку репликация ДНК на ведущей цепи продолжается до тех пор, пока репликация не завершится. Однако вотстающей цепи ДНК-полимераза δ должна постоянно загружаться в начале каждого фрагмента Окадзаки. Это постоянноеинициирование синтеза фрагмента Окадзаки требует повторной загрузки PCNA для эффективной репликации ДНК.
Загрузка PCNA осуществляется комплексом фактора репликации C (RFC). RFC-комплекс состоит из пяти АТФаз: Rfc1, Rfc2, Rfc3, Rfc4 и Rfc5. RFC распознает соединения праймер-матрица и загружает PCNA в эти сайты. Гомотример PCNA открывается RFC гидролизом АТФ, а затемзагружается на ДНК в правильной ориентации, чтобы облегчить ее ассоциацию с полимеразой. Загрузчики зажимов такжемогут выгружать PCNA из ДНК; механизм, необходимый, когда репликация должна быть прекращена.
Репликация ДНК на репликационной вилке может быть остановлена из-за нехватки дезоксинуклеотидтрифосфатов (dNTP) или Повреждение ДНК, приводящее к стрессу репликации. Эта остановка репликации описывается как остановленная репликационная вилка . Белковый комплекс защиты вилки стабилизирует вилку репликации до тех пор, пока небудут устранены повреждения ДНК или другие проблемы репликации. Длительная остановка репликационной вилки может привести к дальнейшему повреждению ДНК. Сигналы остановки отключаются, если устранены проблемы, вызывающие репликационную вилку.
 Изображение теломеразы, постепенно удлиняющей теломерную ДНК.
Изображение теломеразы, постепенно удлиняющей теломерную ДНК. Прекращение репликации эукариотической ДНКтребует различных процессов в зависимости от того, являются ли хромосомы круговой или линейный. В отличие от линейныхмолекул, кольцевые хромосомы способны воспроизводить всю молекулу. Однако две молекулы ДНК останутся связанными вместе. Эта проблема решается декатенацией двух молекул ДНК с помощью топоизомеразы типа II. Топоизомеразы типа II также используются для разделения линейных цепей, поскольку они сложным образом свернуты в нуклеосому внутри клетки.
Как упоминалосьранее, линейные хромосомы сталкиваются с другой проблемой, которая не наблюдается при кольцевой репликации ДНК. Из-за того,что для инициации синтеза ДНК требуется праймер РНК, отстающая цепь находится в невыгодном положении при репликации всей хромосомы. В то время как ведущая цепь может использовать один праймер РНК для удлинения 5'-конца реплицирующейся цепи ДНК, несколько праймеров РНК отвечают за синтез отстающей цепи, создавая фрагменты Окадзаки. Это приводит к проблеме из-за того, чтоДНК-полимераза может присоединяться только к 3'-концу цепи ДНК. 3'-5'-действие ДНК-полимеразы вдоль родительской цепи оставляеткороткую одноцепочечную ДНК (оцДНК) на 3'-конце родительской цепи, когда фрагменты Окадзаки были восстановлены. Поскольку репликация происходит в противоположных направлениях на противоположных концах родительских хромосом, каждая нить является отстающей нитью на одном конце. Со временем это приведет к прогрессивному укорочению обеих дочерних хромосом. Этоизвестно как проблема конца репликации.
Проблема конца репликации решается в эукариотических клетках с помощью областей теломер и теломеразы. Теломеры расширяют 3'-конец родительской хромосомы за 5'-конец дочерней цепи. Эта одноцепочечная структура ДНК может действовать как точка начала репликации, которая рекрутирует теломеразу. Теломераза - это специализированная ДНК-полимераза, которая состоит из нескольких белковых субъединиц и компонента РНК. Компонент РНКтеломеразы отжигается с одноцепочечным 3'-концом матричной ДНК и содержит 1,5 копии теломерной последовательности. Теломеразасодержит субъединицу белка, которая представляет собой обратную транскриптазу , называемую обратной транскриптазой теломеразы или TERT. TERT синтезирует ДНК до конца матричной теломеразной РНК, а затем отключается. Этот процесс можно повторять сколько угодно раз с удлинением 3'-конца родительской молекулы ДНК. Это 3'-добавление обеспечивает матрицу дляудлинения 5'-конца дочерней цепи за счет синтеза ДНК отстающей цепи. Регулирование активности теломеразы осуществляетсятеломер-связывающими белками.
Репликация прокариотической ДНК является двунаправленной; в репликативном ориджине, реплисомные комплексы создаются на каждом конце репликационного ориджина, и реплисомы удаляются друг от друга от начальной начальной точки. У прокариот двунаправленная репликация инициируется в одном репликативном ориджинена кольцевой хромосоме и заканчивается в сайте, противоположном начальному началу ориджина. Эти области терминации имеютпоследовательности ДНК, известные как Ter-сайты. Эти сайты Ter связаны с белком Tus. Комплекс Ter-Tus способен останавливать активность геликазы, прекращая репликацию.
В эукариотических клетках прекращение репликации обычно происходит в результате столкновения двух репликативных вилок между двумя активными источниками репликации. Место столкновения зависит от времениначала стрельбы. Таким образом, если вилка репликации останавливается или разрушается на определенном сайте, репликациясайта может быть восстановлена, когда реплисома, перемещающаяся в противоположном направлении, завершает копирование региона. Существуют запрограммированные барьеры вилки репликации (RFB), связанные с белками RFB в различных местах по всему геному, которые способны останавливать или приостанавливать вилки репликации, останавливая развитие реплисомы.
Было обнаружено, что репликация происходит локализованно в ядре клетки. Вопреки традиционному представлению оперемещении репликационных вилок по застойной ДНК, возникла концепция репликационных фабрик, что означает, что репликационные вилки сосредоточены в некоторых иммобилизованных «фабричных» областях, через которые нити матричной ДНК проходят как конвейерные ленты.
.
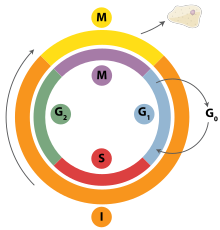 клеточный цикл для эукариотических клеток.
клеточный цикл для эукариотических клеток.Репликация ДНК - это строго организованный процесс, который контролируется в контексте клеточный цикл. Прогресс вклеточном цикле и, в свою очередь, репликация ДНК строго регулируется образованием и активацией пререпликативных комплексов (пре-RC), что достигается за счет активации и инактивации циклин-зависимых киназ (Cdks, CDK). В частности, именно взаимодействия циклинов и циклинзависимых киназ ответственны за переход из G 1 в S-фазу.
Во время фазы G1 клеточного цикла наблюдается низкий уровень активности CDK. Этот низкий уровень активности CDK позволяетформировать новые комплексы пре-RC, но недостаточен для инициирования репликации ДНК вновь образованными пре-RC. Во время остальных фаз клеточного цикла наблюдается повышенный уровень активности CDK. Этот высокий уровень активности CDK отвечает за инициирование репликации ДНК, а также за ингибирование образования нового комплекса пре-RC. Как только репликация ДНК былаинициирована, пре-RC комплекс разрушается. Благодаря тому факту, что уровни CDK остаются высокими во время S-фазы, G 2 иM-фаз клеточного цикла, новые пре-RC-комплексы не могут образовываться. Все это помогает гарантировать, что инициация не может произойти, пока клеточное деление не завершится.
В дополнение к циклинзависимым киназам, новый раунд репликации, как полагают, предотвращается посредством подавления Cdt1. Это достигается за счет деградации Cdt1, а также за счетингибирующего действия белка, известного как геминин. Геминин прочно связывается с Cdt1 и считается основным ингибиторомповторной репликации. Геминин сначала появляется в S-фазе и разрушается при переходе от метафазы к анафазе, возможно, за счет убиквинирования с помощью комплекса, стимулирующего анафазу (APC).
Различные контрольные точки клеточного цикла присутствуют в течение клеточного цикла, что определяет, будет ли клетка полностью продвигаться черезделение. Важно отметить, что при репликации G 1 или ограничение, контрольная точка определяет, начнется ли инициациярепликации или будет ли ячейка помещена в стадию покоя, известную как G0. Клеткам на стадии G 0 клеточного цикла не позволяют инициировать цикл репликации, поскольку поддерживающие белки минихромосомы не экспрессируются. Переход в S-фазу указывает на то, что репликация началась.
Чтобы сохранить генетическуюинформацию во время деления клетки, репликация ДНК должна выполняться с высокой точностью. Для достижения этой задачи вэукариотических клетках в определенные моменты процесса репликации присутствуют белки, которые способны обнаруживать любые ошибки во время репликации ДНК и сохранять целостность генома. Эти белки контрольных точек способны останавливать клеточный цикл от входа в митоз, чтобы дать время на восстановление ДНК. Белки контрольных точек также участвуют в некоторых путях репарацииДНК, в то время как они стабилизируют структуру репликационной вилки, чтобы предотвратить дальнейшее повреждение. Этибелки контрольных точек необходимы для предотвращения передачи мутаций или других хромосомных аберраций потомству.
Эукариотические белки контрольных точек хорошо консервативны и включают две фосфатидилинозитол-3-киназы киназы (PIKKs), ATR и ATM. И ATR, и ATM имеют общую последовательность фосфорилирования-мишени, мотив SQ / TQ, но ихиндивидуальные роли в клетках различаются.
ATR участвует в остановке клеточного цикла в ответ на двухцепочечные разрывыДНК. ATR имеет обязательного партнера по контрольной точке, ATR-взаимодействующий белок (ATRIP), и вместе эти два белка реагируют на участки одноцепочечной ДНК, покрытые репликационным белком A (RPA). Образование одноцепочечной ДНК происходит часто, чаще во время репликационного стресса. ATR-ATRIP способен останавливать клеточный цикл для сохранения целостности генома.ATR обнаруживается на хроматине во время фазы S, подобно RPA и класпину.
Генерация участков одноцепочечной ДНК важнадля инициирования путей контрольных точек ниже повреждения репликации. Как только одноцепочечная ДНК становится достаточно длинной, одноцепочечная ДНК, покрытая RPA, может рекрутировать ATR-ATRIP. Чтобы стать полностью активной, киназа ATR полагается на сенсорные белки, которые определяют, локализованы ли белки контрольной точки в допустимом месте стресса репликации ДНК.Гетеротримерный зажим RAD9 - HUS1 - Rad1 (9-1-1) и его загрузчик зажимов RFC способныраспознавать зазоры или порезанная ДНК. Загрузчик зажимов RFC загружает 9-1-1 на поврежденную ДНК. Присутствия 9-1-1 на ДНК достаточно для облегчения взаимодействия между ATR-ATRIP и группой белков, называемых медиаторами контрольных точек, такими как TOPBP1 и Mrc1 / claspin. TOPBP1 взаимодействует и рекрутирует фосфорилированный компонент Rad9 в 9-1-1 исвязывает ATR-ATRIP, который фосфорилирует Chk1. Mrc1 / Claspin также необходим для полной активации ATR-ATRIP, которыйфосфорилирует Chk1, главную эффекторную киназу нижестоящей контрольной точки. Claspin является компонентом реплисомы и содержит домен для стыковки с Chk1, раскрывая специфическую функцию Claspin во время репликации ДНК: стимулирование передачи сигналов контрольной точки на реплисоме.
Передача сигналов Chk1 жизненно важна для остановки клеточного цикла и предотвращениевхода клеток в митоз с неполной репликацией ДНК или повреждением ДНК. Chk1-зависимое ингибирование Cdk важно для функцииконтрольной точки ATR-Chk1, а также для остановки клеточного цикла и предоставления достаточного времени для завершения механизмов репарации ДНК, что, в свою очередь, предотвращает наследование поврежденной ДНК. Кроме того, Chk1-зависимое ингибирование Cdk играет критическую роль в ингибировании инициирования инициирования во время S фазы. Этот механизм предотвращаетнепрерывный синтез ДНК и необходим для защиты генома в присутствии стресса репликации и потенциальных генотоксических условий.Таким образом, активность ATR-Chk1 дополнительно предотвращает потенциальные проблемы репликации на уровне единичных источников репликации путем ингибирования инициации репликации по всему геному, пока сигнальный каскад, поддерживающий остановку клеточного цикла, не будет отключен.
 Изображение репликации через гистоны. Гистоныудаляются из ДНК комплексом FACT и Asf1. Гистоны повторно собираются на вновь реплицированную ДНК после репликационной вилки спомощью CAF-1 и Rtt106.
Изображение репликации через гистоны. Гистоныудаляются из ДНК комплексом FACT и Asf1. Гистоны повторно собираются на вновь реплицированную ДНК после репликационной вилки спомощью CAF-1 и Rtt106. Эукариотическая ДНК должна быть плотно уплотнена, чтобы соответствовать ограниченному пространству ядра. Хромосомы упаковываются путем обертывания 147 нуклеотидов вокруг октамера белков гистонов, образуя нуклеосому. Октамер нуклеосомы включает по две копии каждого гистона H2A, H2B, H3 и H4. Из-затесной ассоциации гистоновых белков с ДНК, эукариотические клетки имеют белки, которые предназначены для ремоделирования гистоновперед репликационной вилкой, чтобы обеспечить плавное развитие реплисомы. Существуют также белки, участвующие в повторной сборке гистонов позади репликационной вилки для восстановления конформации нуклеосомы.
Есть несколько гистоновых шаперонов, которые, как известно, участвуют в сборке нуклеосом после репликации. Было обнаружено, что комплекс FACT взаимодействует с комплексом ДНК-полимераза-α-примаза, а субъединицы комплекса FACT генетически взаимодействуют с факторамирепликации. Комплекс FACT представляет собой гетеродимер, который не гидролизует АТФ, но способен способствовать «разрыхлению» гистонов в нуклеосомах, но вопрос о том, как комплекс FACT может ослабить тесную ассоциацию гистонов для удаления ДНК, остается без ответа.
Другой шаперон гистонов, который ассоциируется с реплисомой, - это Asf1, которыйвзаимодействует с комплексом Mcm, зависящим от димеров гистонов H3-H4. Asf1 способен передавать вновь синтезированный димер H3-H4 кфакторам отложения за вилкой репликации, и эта активность делает димеры гистона H3-H4 доступными в месте отложения гистона сразу после репликации. Asf1 (и его партнер Rtt109) также участвует в ингибировании экспрессии генов реплицированных генов во время S-фазы.
Гетеротримерный шаперонный фактор сборки хроматина 1 (CAF-1 ) представляет собойхроматин белок образования, который участвует в депонировании гистонов на обеих вновь реплицированных цепях ДНК с образованием хроматина.CAF-1 содержит мотив связывания PCNA, называемый PIP-боксом, который позволяет CAF-1 связываться с реплисомой через PCNA и способен депонировать димеры гистона H3-H4 на вновь синтезированную ДНК. Шаперон Rtt106 также участвует в этом процессе и ассоциируется с димерами CAF-1 и H3-H4 во время образования хроматина. Эти процессы загружают вновь синтезированныегистоны в ДНК.
После отложения гистонов H3-H4 нуклеосомы образуются за счет ассоциации гистона H2A-H2B. Считается, что этотпроцесс происходит через комплекс FACT, поскольку он уже связан с реплисомой и способен связывать свободные H2A-H2B, или существует возможность другого шаперона H2A-H2B, Nap1. Электронно-микроскопические исследования показывают, что это происходит очень быстро, так как можно наблюдать образование нуклеосом, образующих всего несколько сотен пар оснований послерепликационной вилки. Следовательно, весь процесс формирования новых нуклеосом происходит сразу после репликации из-за связываниягистоновых шаперонов с реплисомой.
По сравнению с репликацией прокариотической ДНК завершение репликации эукариотической ДНК является более сложным и включает несколько точек начала репликации и репликативные белки, которые необходимо выполнить. Прокариотическая ДНК имееткруглую форму и имеет только одну точку начала репликации, когда репликация начинается. Напротив, ДНК эукариот линейна. Прирепликации существует до тысячи источников репликации.
Эукариотическая ДНК двунаправлена. Здесь значение слова «двунаправленный» иное. Линейная ДНК эукариот имеет множество источников (называемых O) и концов (называемых T). "T" присутствует справа от "O". Один «О» и один «Т» вместе образуют один репликон. После образования комплекса пре-инициации, когда одинрепликон начинает элонгацию, инициация начинается во втором репликоне. Теперь, если первый репликон движется по часовой стрелке,второй репликон движется против часовой стрелки, пока не будет достигнута «Т» первого репликона. В точке «T» оба репликона сливаются, чтобы завершить процесс репликации. Между тем, второй репликон также движется в прямом направлении, чтобы встретиться с третьим репликоном. Это движение двух репликонов по часовой стрелке и против часовой стрелки называетсядвунаправленной репликацией.
Репликация эукариотической ДНК требует точной координации всех ДНК-полимераз и связанных белков длярепликации всего генома каждый раз при делении клетки. Этот процесс достигается с помощью серии этапов сборки белков в точках начала репликации, в основном фокусируясь на регуляции репликации ДНК на ассоциации геликазы MCM с ДНК. Эти источники репликации определяют количество белковых комплексов, которые будут образовываться для инициации репликации. Впрокариотической ДНК регуляция репликации сосредоточена на связывании белка инициатора DnaA с ДНК, при этом инициация репликации происходитнесколько раз в течение одного клеточного цикла. Как прокариотическая, так и эукариотическая ДНК используют связывание и гидролиз АТФ для непосредственной загрузки геликазы, и в обоих случаях геликаза загружается в неактивной форме. Однако эукариотические геликазы представляют собой двойные гексамеры, которые загружены на двухцепочечную ДНК, тогда какпрокариотические геликазы представляют собой одноцепочечные гексамеры, загруженные в одноцепочечную ДНК.
Сегрегация хромосом - еще одноразличие между прокариотическими и эукариотическими клетками. Быстро делящиеся клетки, такие как бактерии, часто начинают отделять хромосомы, которые все еще находятся в процессе репликации. В эукариотических клетках сегрегация хромосом в дочерние клетки не начинается до тех пор, пока репликация не завершится во всех хромосомах. Однако, несмотря на этиразличия, основной процесс репликации одинаков как для прокариотической, так и для эукариотической ДНК.
| Репликация прокариотической ДНК | Репликация эукариотической ДНК |
|---|---|
| Происходит внутри цитоплазмы | Происходит внутри ядра |
| Только одна точка начала репликации на молекулу ДНК | Имеют множество источников репликации в каждой хромосоме |
| Начало репликации составляет примерно 100-200 или более нуклеотидов в длину | Каждая точка начала репликации состоитиз примерно 150 нуклеотидов |
| Репликация происходит в одной точке в каждой хромосома | Репликация происходит в нескольких точкаходновременно в каждой хромосоме |
| Имеется только одна точка начала репликации | Имеется несколько точек начала репликации |
| Инициирование осуществляется белками DnaA и DnaB | Инициирование осуществляется комплексом распознавания происхождения |
| Требуется топоизомераза | Требуется топоизомераза |
| Репликация очень быстрая | Репликация очень медленная |
Список основных белков, участвующих в репликацииэукариотической ДНК:
| Белок | Функция в репликации эукариотической ДНК |
|---|---|
| AND1 | Загружает ДНК-полимеразу α на хроматин вместе с комплексом CMG на отстающей цепи. Также известен как Ctf4 у почкующихся дрожжей. |
| Cdc45 | Требуется для стадий инициации и удлинения репликации ДНК. Входит в состав геликазного комплекса Mcm2-7.Требуется после этапа перед RC для загрузки различных белков для инициации и удлинения. |
| Комплекс Cdc45-Mcm-GINS (CMG) | ФункциональнаяДНК-геликаза в эукариотических клетках |
| Cdc6 | Требуется для сборки комплекса Mcm2-7 в ORC в сочетании с Cdt1. |
| Киназа Cdc7-Dbf4 или Dbf4-зависимая киназа (DDK) | Протеинкиназа, необходимая для инициации репликации ДНК, вероятно, посредством фосфорилирования поддерживающих белков минихромосомы. |
| Cdt1 | Загружает комплекс Mcm 2-7 на ДНК в ORC на этапе до RC / лицензирования. У многоклеточных животных подавляется близнецами. |
| Claspin | Соедините синтез ведущейцепи с геликазной активностью комплекса CMG. Работает с Mrc1 |
| Ctf4 | Загружает ДНК-полимеразу α на хроматин вместе с комплексом CMG на отстающей цепи. Гомолог многоклеточных животных известен как AND-1. |
| Циклин-зависимая киназа (CDK) | Циклин-зависимая протеинкиназа, необходимая для инициации репликации и для других последующихстадий. |
| Dna2 | 5'-эндонуклеаза лоскута и геликаза, участвующие в процессинге фрагментов Окадзаки. |
| ДНК-лигаза I | Соединяетфрагменты Окадзаки во время репликации ДНК. Лигазная активность также необходима для репарации ДНК и рекомбинации. |
| ДНК-полимераза α (Pol α) | Обладает активностью примазы, которая необходима для инициации синтеза ДНК как на ведущей, так и на отстающей цепи. |
| ДНК-полимераза δ (Pol δ) | Требуется для завершения синтезафрагментов Окадзаки на отстающей цепи, которые были инициированы ДНК-полимеразой α. |
| ДНК-полимераза ε (Pol ε) | Полимераза ведущей цепи.Синтезирует ДНК на вилке репликации. Связывается на ранней стадии происхождения через Dbp11 и необходимо для загрузки ДНК-полимеразы α. |
| Dpb11 | Белок инициации репликации ДНК. Загружает ДНК-полимеразу ε на пререпликационные комплексы в источниках. |
| Fen1 | 5'-эндонуклеаза лоскута, участвующая в процессинге фрагментов Окадзаки. |
| Геминин | Белок, обнаруженный у многоклеточных животных и отсутствующий у дрожжей. Связывается с Cdt1 и инактивирует его, тем самым регулируяобразование пререпликативного / инициирующего комплекса. Также предлагается способствовать образованию пре-RC путем связывания и, таким образом, предотвращения деградации Cdt1 |
| GINS | Тетрамерный комплекс, состоящий из Sld5, Psf1, Psf2, Psf3. Связывается с пререпликативным комплексом во время инициации и перемещается с вилками репликации наэтапе элонгации. Требуется для стадии элонгации репликации ДНК и, возможно, является частью геликазного комплекса Mcm. |
| Поддерживающиеминихромосомы белки (Mcm) | Шесть различных белков семейства AAA + ATPase, которые образуют гексамер в растворе. Этот гексамер рекрутируется и загружается ORC, Cdc6 и Cdt1 и образует двойной гексамер, который топологически связан вокруг ДНК с образованием солеустойчивого пререпликативного комплекса. При инициации репликации Mcm2-7 удаляется от ORCс помощью вилки репликации. |
| Mcm10 | Требуется для стадий инициации и удлинения репликации ДНК. Участвует в связывании хроматинаCdc45 и ДНК-полимеразы α. Также требуется для стабильности каталитической субъединицы ДНК-полимеразы α в почкующихся дрожжах S. cerevisiae. |
| Mrc1 | Соедините синтез ведущей цепи с геликазной активностью комплекса CMG. Гомолог Metazoan известен как Claspin. |
| Комплекс распознавания источника (ORC) | Гетерогексамерный комплекс, состоящий избелков Orc1 – Orc6. Связывается с ДНК и собирает комплекс Mcm2-7 на хроматин вместе с Cdc6 и Cdt1. |
| Ядерный антиген пролиферирующихклеток (PCNA) | Тримерный белок с кольцевой структурой, содержащий ДНК, предотвращающую диссоциацию ДНК-полимеразы. Действует как скользящий зажим для полимераз δ и ε, тем самым улучшая процессивность репликативных полимераз. |
| Фактор репликации C (RFC) | Загружает PCNA на примированные матрицы и участвует в переключении междуДНК-полимеразой a и репликативными полимеразами δ и ε. |
| Барьеры репликационной вилки (RFB) | Связаны RFB-белками в различных местах повсему геному. Умеют останавливать или приостанавливать вилки репликации, останавливая развитие реплисомы. |
| Репликационный белок A (RPA) | Гетеротримерный одноцепочечный связывающий белок. Стабилизирует одноцепочечную ДНК на вилке репликации. |
| РНКаза H | Рибонуклеаза, которая расщепляет РНК, гибридизованную с ДНК. Участвует вобработке фрагментов Окадзаки. |
| Sld2 | Функции инициации репликации. Ключевой субстрат CDK, фосфорилирование способствует взаимодействию сDpb11. Требуется для запуска репликации. |
| Sld3 | Функции инициации репликации. Ключевой субстрат CDK, фосфорилирование способствует взаимодействию с Dpb11. Требуется для запуска репликации. |
| Теломераза | Рибонуклеопротеин, который добавляет последовательность ДНК «TTAGGG», повторяется на 3'-конце цепей ДНК в теломерах. |
| Топоизомеразы | Регулируют перекручивание или обратное скручивание ДНК |