
Войти
| рибонуклеаза H | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Кристаллографическая структура РНКазы HI E. coli. Кристаллографическая структура РНКазы HI E. coli. | |||||||||
| Идентификаторы | |||||||||
| ЕС нет. | 3.1.26.4 | ||||||||
| № CAS | 9050-76-4 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| Структуры PDB | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | Amigo / QuickGO | ||||||||
| |||||||||
| ретровирусная рибонуклеаза H | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| ЕС нет. | 3.1.26.13 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| Структуры PDB | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Рибонуклеазы Н (сокращенно РНКазы Н или RNH) представляет собой семейство не- последовательность -специфический эндонуклеазы ферменты, которые катализируют расщепление РНК в РНК / ДНК - субстрата посредством гидролитического механизма. Члены семейства РНКазы H можно найти почти у всех организмов, от бактерий до архей и эукариот.
Семейство разделено на эволюционно родственные группы с немного разными предпочтениями субстратов, в широком смысле обозначаемые рибонуклеазами H1 и H2. Геном человека кодирует как H1 и H2. Человеческая рибонуклеаза H2 представляет собой гетеротримерный комплекс, состоящий из трех субъединиц, мутации в любой из которых являются одной из генетических причин редкого заболевания, известного как синдром Айкарди – Гутьера. Третий тип, тесно связанный с H2, встречается только у нескольких прокариот, тогда как H1 и H2 встречаются во всех сферах жизни. Кроме того, РНКаза H1-подобные домены ретровирусной рибонуклеазы H встречаются в многодоменных белках обратной транскриптазы, которые кодируются ретровирусами, такими как ВИЧ, и необходимы для репликации вируса.
В эукариот, рибонуклеазы H1 участвует в репликации ДНК из митохондриального генома. И H1, и H2 участвуют в задачах поддержания генома, таких как обработка структур R-петли.
Рибонуклеазы Н представляет собой семейство эндонуклеаз ферментов с общей подложки специфичности для РНК - нити РНК - ДНК - дуплексов. По определению, РНКазы H расщепляют фосфодиэфирные связи основной цепи РНК, оставляя 3'- гидроксильную и 5'- фосфатную группы. РНКазы H были предложены как члены эволюционно родственного суперсемейства, включающего другие нуклеазы и ферменты, обрабатывающие нуклеиновые кислоты, такие как ретровирусные интегразы, ДНК- транспозазы, резольвазы соединений Холлидея, белки Piwi и Argonaute, различные экзонуклеазы и сплайсосомный белок Prp8.
РНКазы H можно в общих чертах разделить на два подтипа, H1 и H2, которым по историческим причинам даны арабские цифровые обозначения у эукариот и римские цифровые обозначения у прокариот. Таким образом, кишечная палочка РНКазы HI является гомологом человека разумного РНКазы H1. У E. coli и многих других прокариот ген rnhA кодирует HI, а ген rnhB кодирует HII. Третий родственный класс, называемый HIII, встречается у некоторых бактерий и архей ; он тесно связан с прокариотическими ферментами HII.
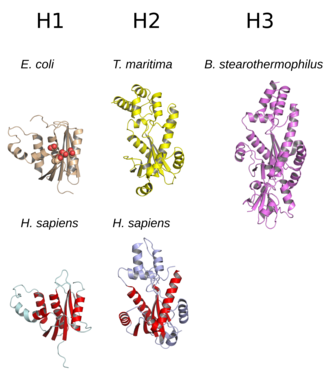 Сравнение структур репрезентативных белков рибонуклеазы H каждого подтипа. В белке E. coli (бежевый, вверху слева) четыре консервативных остатка активного сайта показаны сферами. В белках H. sapiens структурное ядро, общее между подтипами H1 и H2, показано красным. Структуры визуализируются из: E. coli, PDB : 2RN2 ; T. maritima, PDB : 303F ; B. stearothermophilus, PDB : 2D0B ; H. sapiens H1, PDB : 2QK9 ; H. sapiens, PDB : 3P56.
Сравнение структур репрезентативных белков рибонуклеазы H каждого подтипа. В белке E. coli (бежевый, вверху слева) четыре консервативных остатка активного сайта показаны сферами. В белках H. sapiens структурное ядро, общее между подтипами H1 и H2, показано красным. Структуры визуализируются из: E. coli, PDB : 2RN2 ; T. maritima, PDB : 303F ; B. stearothermophilus, PDB : 2D0B ; H. sapiens H1, PDB : 2QK9 ; H. sapiens, PDB : 3P56. Структура РНКазы Н, обычно состоит из 5- ти многожильных бета-листа, окруженного распределением альфа-спирали. Все РНКазы H имеют активный сайт, сосредоточенный на консервативном мотиве последовательности, состоящем из остатков аспартата и глутамата, часто называемом мотивом DEDD. Эти остатки взаимодействуют с каталитически необходимыми ионами магния.
РНКазы H2 больше, чем H1, и обычно имеют дополнительные спирали. Домена организация ферментов изменяется; некоторые прокариотические и большинство эукариотических членов группы H1 имеют дополнительный небольшой домен на N-конце, известный как «гибридный связывающий домен», который облегчает связывание с гибридными дуплексами РНК: ДНК и иногда обеспечивает повышенную процессивность. В то время как все члены группы H1 и прокариотические члены группы H2 действуют как мономеры, ферменты H2 эукариот являются облигатными гетеротримерами. Прокариотические ферменты HIII являются членами более широкой группы H2 и имеют общие структурные особенности с H2 с добавлением N-концевого связывающего домена TATA-бокса. Домены ретровирусной РНКазы H, встречающиеся в мультидоменных белках обратной транскриптазы, имеют структуру, очень напоминающую группу H1.
РНКазы H1 были тщательно изучены для изучения взаимосвязи между структурой и ферментативной активностью. Они также используются, особенно гомолог E. coli, в качестве модельных систем для изучения сворачивания белков. Внутри группы H1 была выявлена взаимосвязь между более высокой аффинностью связывания субстрата и наличием структурных элементов, состоящих из спирали и гибкой петли, обеспечивающих большую и более основную поверхность связывания субстрата. C-спираль имеет разбросанное таксономическое распределение; он присутствует в гомологах E. coli и человеческой РНКазы H1 и отсутствует в домене РНКазы H ВИЧ, но примеры ретровирусных доменов с С-спиралями действительно существуют.
Ферменты рибонуклеазы Н расщепляют фосфодиэфирные связи РНК в двухцепочечном гибриде РНК: ДНК, оставляя 3'- гидроксильную и 5'- фосфатную группы на обоих концах участка разреза с механизмом катализа с двумя ионами металлов, в котором две двухвалентные катионы, такие как Mg2 + и Mn2 +, непосредственно участвуют в каталитической функции. В зависимости от различий в их аминокислотных последовательностях эти РНКазы H классифицируются на РНКазы 1 и 2 типа H. РНКазы H типа 1 содержат прокариотические и эукариотические РНКазы H1 и ретровирусные РНКазы H. РНКазы H типа 2 содержат прокариотические и эукариотические РНКазы H2 и бактериальная РНКаза H3. Эти РНКазы H существуют в мономерной форме, за исключением эукариотических РНКаз H2, которые существуют в гетеротримерной форме. РНКазы H1 и H2 имеют разные предпочтения в отношении субстратов и разные, но частично совпадающие функции в клетке. У прокариот и низших эукариот ни один из ферментов не важен, тогда как оба считаются важными для высших эукариот. Комбинированная активность ферментов H1 и H2 связана с поддержанием стабильности генома из-за деградации ферментами РНК-компонента R-петель.
| Идентификаторы | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Условное обозначение | РНКаза H | ||||||||
| Pfam | PF00075 | ||||||||
| Клан пфам | CL0219 | ||||||||
| ИнтерПро | IPR002156 | ||||||||
| ПРОФИЛЬ | PS50879 | ||||||||
| |||||||||
Ферменты рибонуклеазы H1 требуют по меньшей мере четырех пар оснований, содержащих рибонуклеотиды, в субстрате и не могут удалить один рибонуклеотид из цепи, которая в противном случае состоит из дезоксирибонуклеотидов. По этой причине маловероятно, что ферменты РНКазы H1 участвуют в процессинге праймеров РНК из фрагментов Окадзаки во время репликации ДНК. РНКаза H1 не важна для одноклеточных организмов, где она была исследована; в E. coli нокауты РНКазы H1 придают чувствительный к температуре фенотип, а у S. cerevisiae они вызывают дефекты стрессовой реакции.
У многих эукариот, включая млекопитающих, гены РНКазы H1 включают митохондриальную нацеливающую последовательность, приводящую к экспрессии изоформ с присутствием MTS и без нее. В результате РНКаза H1 локализуется как в митохондриях, так и в ядре. В моделях мышей с нокаутом нокаутные мутанты по РНКазе H1 являются летальными во время эмбриогенеза из-за дефектов репликации митохондриальной ДНК. Дефекты репликации митохондриальной ДНК, вызванные потерей РНКазы H1, вероятно, связаны с дефектами процессинга R-петли.
| Идентификаторы | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Условное обозначение | РНКаза HII | ||||||||
| Pfam | PF01351 | ||||||||
| Клан пфам | CL0219 | ||||||||
| ИнтерПро | IPR024567 | ||||||||
| |||||||||
У прокариот РНКаза Н2 ферментативно активна как мономерный белок. У эукариот это облигатный гетеротример, состоящий из каталитической субъединицы A и структурных субъединиц B и C. В то время как субъединица A близко гомологична прокариотической РНКазе H2, субъединицы B и C не имеют явных гомологов у прокариот и плохо сохраняются в последовательность уровня даже среди эукариот. Субъединица B опосредует белок-белковые взаимодействия между комплексом H2 и PCNA, который локализует H2 в фокусах репликации.
Как прокариотические, так и эукариотические ферменты H2 могут расщеплять отдельные рибонуклеотиды в цепи. однако у них несколько разные модели расщепления и предпочтения субстратов: прокариотические ферменты имеют более низкую процессивность и гидролизуют последовательные рибонуклеотиды более эффективно, чем рибонуклеотиды с 5'- дезоксирибонуклеотидом, в то время как эукариотические ферменты более процессивны и гидролизуют оба типа субстрата с одинаковой эффективностью. Субстратная специфичность РНКазы H2 придает ей роль в эксцизионной репарации рибонуклеотидов, удаляя неправильно инкорпорированные рибонуклеотиды из ДНК, в дополнение к процессингу R-петли. Хотя и H1, и H2 присутствуют в ядре клетки млекопитающих, H2 является доминирующим источником активности РНКазы H и важен для поддержания стабильности генома.
Некоторые прокариоты обладают дополнительным геном Н2-типа, обозначенным РНКазой HIII в римско-цифровой номенклатуре, используемой для генов прокариот. Белки HIII более близки к группе H2 по идентичности последовательностей и структурному сходству, но имеют предпочтения в отношении субстратов, которые больше напоминают H1. В отличие от HI и HII, которые широко распространены среди прокариот, HIII обнаруживается только у нескольких организмов с разбросанным таксономическим распределением; он несколько чаще встречается у архей и редко или никогда не встречается в том же прокариотическом геноме, что и HI.
 Механизм реакции для катализа РНКазы H с использованием двух ионов металлов в домене РНКазы H ВИЧ-1
Механизм реакции для катализа РНКазы H с использованием двух ионов металлов в домене РНКазы H ВИЧ-1 Активный сайт почти всех РНКазы Н содержит четыре отрицательно заряженных аминокислотных остатков, известных как DEDD мотив; часто также присутствует гистидин, например, в ВИЧ-1, человеке или E. coli.
Заряженные остатки связывают два иона металла, необходимые для катализа; в физиологических условиях это ионы магния, но марганец также обычно поддерживает ферментативную активность, тогда как кальций или высокая концентрация Mg2 + подавляет активность.
Основываясь на экспериментальных данных и компьютерном моделировании, фермент активирует молекулу воды, связанную с одним из ионов металла, с помощью консервативного гистидина. Переходное состояние является ассоциативным по природе и образует промежуточное соединение с уходящей группой протонированного фосфата и депротонированного алкоксида. Уходящая группа протонируется через глутамат, который имеет повышенное значение pKa и, вероятно, протонируется. Механизм подобен РНКазе Т и субъединице RuvC в ферменте Cas9, которые также используют гистидиновый и двухметаллический ионы.
Механизм высвобождения расщепленного продукта до сих пор не выяснен. Экспериментальные данные кристаллографии с временным разрешением и аналогичных нуклеаз указывают на роль третьего иона в реакции, задействованной в активном центре.
Геном человека содержит четыре генов, кодирующих РНКазы H:
Кроме того, генетический материал ретровирусного происхождения часто появляется в геноме, что отражает интеграцию геномов эндогенных ретровирусов человека. Такие события интеграции приводят к присутствию генов, кодирующих обратную транскриптазу ретровируса, которая включает домен РНКазы Н. Пример - ERVK6. Длинный концевой повтор (LTR) и не длинный концевой повтор (без LTR) ретротранспозоны также распространены в геноме и часто включают в свои собственные домены РНКазы H, со сложной эволюционной истории.
 Структура тримерного комплекса H2 человека: каталитическая субъединица A выделена синим, структурная субъединица B - коричневым, а структурная субъединица C - розовым. Хотя субъединицы B и C не взаимодействуют с активным сайтом, они необходимы для активности. Каталитические остатки в активном центре показаны пурпурным цветом. Позиции, показанные желтым, соответствуют известным мутациям AGS. Наиболее распространенная мутация AGS - аланин на треонин в положении 177 субъединицы B - показана зеленой сферой. Многие из этих мутаций не нарушают каталитическую активность in vitro, но дестабилизируют комплекс или препятствуют белок-белковым взаимодействиям с другими белками в клетке.
Структура тримерного комплекса H2 человека: каталитическая субъединица A выделена синим, структурная субъединица B - коричневым, а структурная субъединица C - розовым. Хотя субъединицы B и C не взаимодействуют с активным сайтом, они необходимы для активности. Каталитические остатки в активном центре показаны пурпурным цветом. Позиции, показанные желтым, соответствуют известным мутациям AGS. Наиболее распространенная мутация AGS - аланин на треонин в положении 177 субъединицы B - показана зеленой сферой. Многие из этих мутаций не нарушают каталитическую активность in vitro, но дестабилизируют комплекс или препятствуют белок-белковым взаимодействиям с другими белками в клетке. В небольших исследованиях мутации в человеческой РНКазе H1 были связаны с хронической прогрессирующей внешней офтальмоплегией, общим признаком митохондриальных заболеваний.
Мутации в любой из трех субъединиц РНКазы Н2 хорошо известны как причины редкого генетического заболевания, известного как синдром Айкарди – Гутьера (AGS), которое проявляется в виде неврологических и дерматологических симптомов в раннем возрасте. Симптомы AGS очень напоминают симптомы врожденной вирусной инфекции и связаны с несоответствующей активацией интерферона I типа. AGS также может быть вызван мутациями в других генах: TREX1, SAMHD1, ADAR и MDA5 / IFIH1, каждый из которых участвует в процессинге нуклеиновой кислоты. При характеристике распределения мутаций в популяции пациентов с AGS было обнаружено 5% всех мутаций AGS в RNASEH2A, 36% в 2B и 12% в 2C. Мутации в 2B были связаны с несколько более легкими неврологическими нарушениями и с отсутствием индуцированной интерфероном активации генов, которая может быть обнаружена у пациентов с другими генотипами, связанными с AGS.
 Кристаллическая структура гетеродимера обратной транскриптазы ВИЧ (желтый и зеленый) с доменом РНКазы H, показанным синим цветом (активный центр в пурпурных сферах). Оранжевая цепь нуклеиновой кислоты - это РНК, красная цепь - это ДНК.
Кристаллическая структура гетеродимера обратной транскриптазы ВИЧ (желтый и зеленый) с доменом РНКазы H, показанным синим цветом (активный центр в пурпурных сферах). Оранжевая цепь нуклеиновой кислоты - это РНК, красная цепь - это ДНК. Две группы вирусов используют обратную транскрипцию как часть своего жизненного цикла: ретровирусы, которые кодируют свои геномы в одноцепочечной РНК и реплицируются через промежуточную двухцепочечную ДНК; и вирусы дцДНК-ОТ, которые реплицируют свои геномы двухцепочечной ДНК через промежуточный "прегеном" РНК. Патогенные примеры включают вирус иммунодефицита человека и вирус гепатита B соответственно. Оба кодируют большие многофункциональные белки обратной транскриптазы (RT), содержащие домены РНКазы Н.
Ретровирусные белки RT из вируса ВИЧ-1 и мышиного лейкоза являются наиболее изученными членами этого семейства. Ретровирусная ОТ отвечает за преобразование генома одноцепочечной РНК вируса в двухцепочечную ДНК. Этот процесс требует трех этапов: во-первых, РНК-зависимая активность ДНК-полимеразы производит минус-цепь ДНК из матрицы плюс-цепи РНК, генерируя промежуточный гибрид РНК: ДНК; во-вторых, цепь РНК разрушается; и в-третьих, ДНК-зависимая активность ДНК-полимеразы синтезирует ДНК с положительной цепью, образуя двухцепочечную ДНК в качестве конечного продукта. Второй этап этого процесса осуществляется доменом РНКазы H, расположенным на С-конце белка RT.
РНКаза H выполняет три типа расщепляющих действий: неспецифическая деградация генома плюс-цепи РНК, специфическое удаление праймера тРНК минус-цепи и удаление праймера богатого пуринами полипуринового тракта (PPT) плюсовой цепи. РНКаза H играет роль в праймировании плюс-цепи, но не в обычном способе синтеза новой последовательности праймера. Скорее, РНКаза H создает «праймер» из PPT, который устойчив к расщеплению РНКазой H. Удалив все основания, кроме PPT, PPT используется в качестве маркера для конца области U3 его длинного концевого повтора.
Поскольку активность РНКазы H необходима для вирусной пролиферации, этот домен считается лекарственной мишенью для разработки антиретровирусных препаратов, используемых для лечения ВИЧ / СПИДа и других состояний, вызванных ретровирусами. Идентифицированы ингибиторы ретровирусной РНКазы H нескольких различных хемотипов, многие из которых имеют механизм действия, основанный на хелатировании катионов активного центра. Ингибиторы обратной транскриптазы, которые специфически ингибируют функцию полимеразы RT, широко используются в клинической практике, но не ингибиторы функции РНКазы H; это единственная ферментативная функция, кодируемая ВИЧ, на которую еще не нацелены лекарства, применяемые в клинической практике.
РНКазы H широко распространены и встречаются во всех сферах жизни. Это семейство принадлежит к большему суперсемейству нуклеазных ферментов и считается эволюционно древним. В прокариотических геномах часто присутствуют множественные гены РНКазы H, но существует небольшая корреляция между появлением генов HI, HII и HIII и общими филогенетическими отношениями, предполагая, что горизонтальный перенос генов мог играть роль в установлении распределения этих ферментов. РНКазы HI и HIII редко или никогда не появляются в одном геноме прокариот. Когда геном организма содержит более одного гена РНКазы H, они иногда имеют значительные различия в уровне активности. Было высказано предположение, что эти наблюдения отражают эволюционный паттерн, который сводит к минимуму функциональную избыточность среди генов РНКазы Н. РНКаза HIII, уникальная для прокариот, имеет разбросанное таксономическое распределение и обнаруживается как у бактерий, так и у архей ; считается, что он довольно рано отошел от HII.
Эволюционная траектория РНКазы H2 у эукариот, особенно механизм, с помощью которого эукариотические гомологи стали облигатными гетеротримерами, неясен; субъединицы B и C не имеют явных гомологов у прокариот.
Поскольку РНКаза H специфически разрушает только РНК в двухцепочечных гибридах РНК: ДНК, она обычно используется в качестве лабораторного реагента в молекулярной биологии. Очищенные препараты РНКазы HI и HII E.coli коммерчески доступны. РНКаза HI часто используется для разрушения матрицы РНК после синтеза первой цепи комплементарной ДНК (кДНК) путем обратной транскрипции. Его также можно использовать для расщепления определенных последовательностей РНК в присутствии коротких комплементарных сегментов ДНК. Для обнаружения могут использоваться высокочувствительные методы, такие как поверхностный плазмонный резонанс. РНКазу HII можно использовать для разрушения компонента праймера РНК фрагмента Окадзаки или для введения одноцепочечных разрывов в положения, содержащие рибонуклеотид. Вариант горячего старта ПЦР, известный как РНКазы Н-зависимый ПЦР или rhPCR, был описан с использованием термостабильной РНКазы HII из гипертермофильного archaeon Pyrococcus abyssi. Следует отметить, что белок- ингибитор рибонуклеазы, обычно используемый в качестве реагента, не эффективен при ингибировании активности HI или HII.
Рибонуклеазы H были впервые обнаружены в лаборатории Питера Хаузена, когда исследователи обнаружили активность эндонуклеаз гибридной РНК: ДНК в тимусе теленка в 1969 году и дали ей название «рибонуклеаза H », чтобы обозначить ее гибридную специфичность. Активность РНКазы H была впоследствии обнаружена в E. coli и в образце онковирусов с геномами РНК во время ранних исследований обратной транскрипции вирусов. Позже выяснилось, что экстракт тимуса теленка содержал более одного белка с активностью РНКазы Н и что E. coli содержала два гена РНКазы Н. Первоначально фермент, теперь известный как РНКаза Н2 у эукариот, был обозначен как H1 и наоборот, но названия эукариотических ферментов были изменены, чтобы соответствовать названиям в E. coli, чтобы облегчить сравнительный анализ, в результате чего получена современная номенклатура, в которой обозначены прокариотические ферменты. римскими цифрами и эукариотическими ферментами арабскими цифрами. Прокариотическая РНКаза HIII, о которой сообщалось в 1999 году, была последним идентифицированным подтипом РНКазы H.
Характеристика эукариотической РНКазы Н2 исторически была сложной задачей, отчасти из-за ее низкого содержания. Тщательные усилия по очистке фермента показали, что, в отличие от РНКазы H2 E. coli, эукариотический фермент имеет несколько субъединиц. S.cerevisiae, гомолог E.coli, белка (то есть, Н2А субъединица) было легко идентифицировать с помощью биоинформатики, когда дрожжи геном был секвенирован, но был найден соответствующий белок не иметь ферментативную активность в изоляции. В конце концов, дрожжевые субъединицы B и C были выделены совместной очисткой, и было обнаружено, что они необходимы для ферментативной активности. Однако субъединицы B и C дрожжей имеют очень низкую идентичность последовательностей со своими гомологами в других организмах, и соответствующие человеческие белки были окончательно идентифицированы только после того, как мутации во всех трех оказались причиной синдрома Айкарди-Гутьера.