
Войти
| Калиевый канал внутреннего выпрямителя | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Кристаллическая структура калиевого канала внутреннего выпрямителя Кристаллическая структура калиевого канала внутреннего выпрямителя | |||||||||
| Идентификаторы | |||||||||
| Символ | IRK | ||||||||
| Pfam | PF01007 | ||||||||
| Pfam клан | CL0030 | ||||||||
| InterPro | IPR013521 | ||||||||
| SCOPe | 1n9p / SUPFAM | ||||||||
| TCDB | 1.A.2 | ||||||||
| суперсемейство OPM | 8 | ||||||||
| белок OPM | 3SPG | ||||||||
| |||||||||
Калиевые каналы внутреннего выпрямителя (Kir, IRK ) представляют собой специфическое липидно-зависимое подмножество калиевых каналов. На сегодняшний день идентифицировано семь подсемейств в различных типах клеток млекопитающих, растений и бактерий. Они активируются фосфатидилинозитол-4,5-бисфосфатом (PIP 2 ). Неисправность каналов связана с несколькими заболеваниями. Каналы IRK имеют поровый домен, гомологичный таковому потенциал-управляемых ионных каналов, и фланкирующий трансмембранные сегменты (TMS). Они могут существовать в мембране в виде гомо- или гетероолигомеров, и каждый мономер содержит от 2 до 4 ТМС. С точки зрения функции, эти белки транспортируют калий (K) с большей тенденцией к поглощению K, чем к экспорту K. Процесс внутренней ректификации был обнаружен Денисом Ноблом в клетках сердечной мышцы в 1960-х годах и Ричардом Адрианом и Аланом Ходжкином в 1970 году в клетках скелетных мышц. 331>Содержание
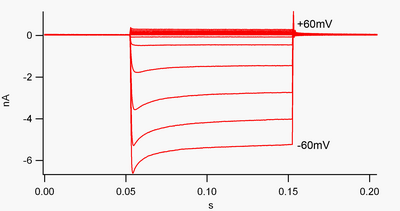 Рисунок 1. Запись тока всей ячейки K ir 2, выпрямляющего внутрь калиевые каналы, экспрессируемые в клетке HEK293. (Это сильно выпрямляющий ток внутрь. Отклонения вниз - это внутренние токи, отклонения вверх - наружные токи, а ось x - время в секундах.) На этом изображении наложено 13 ответов. Самая нижняя кривая - это ток, вызванная скачком напряжения до -60 мВ, а самая верхняя - до +60 мВ относительно потенциала покоя, что близко к потенциалу обращения K в этой экспериментальной системе. Остальные кривые расположены с шагом 10 мВ между ними.
Рисунок 1. Запись тока всей ячейки K ir 2, выпрямляющего внутрь калиевые каналы, экспрессируемые в клетке HEK293. (Это сильно выпрямляющий ток внутрь. Отклонения вниз - это внутренние токи, отклонения вверх - наружные токи, а ось x - время в секундах.) На этом изображении наложено 13 ответов. Самая нижняя кривая - это ток, вызванная скачком напряжения до -60 мВ, а самая верхняя - до +60 мВ относительно потенциала покоя, что близко к потенциалу обращения K в этой экспериментальной системе. Остальные кривые расположены с шагом 10 мВ между ними. Канал, который "выпрямляется вовнутрь", - это канал, который пропускает ток (положительный заряд) легче во внутреннем направлении (в ячейку), чем во внешнем направлении (из клетка). Считается, что этот ток может играть важную роль в регуляции активности нейронов, помогая стабилизировать мембранный потенциал покоя клетки.
По соглашению, входящий ток (положительный заряд, движущийся в ячейку) отображается в фиксаторах напряжения как отклонение вниз, в то время как наружный ток (положительный заряд движется из ячейки). как отклонение вверх. При мембранных потенциалах, отрицательных по отношению к обратному потенциалу калия, внутренние выпрямляющие K-каналы поддерживают поток положительно заряженных ионов K в клетку, подталкивая мембранный потенциал обратно к потенциалу покоя. Это можно увидеть на рисунке 1: когда мембранный потенциал ограничивается отрицательным значением потенциала покоя канала (например, -60 мВ), течет внутренний ток (т.е. положительный заряд течет в ячейку). Однако, когда мембранный потенциал установлен положительным по отношению к потенциалу покоя канала (например, +60 мВ), эти каналы пропускают очень небольшой ток. Проще говоря, этот канал пропускает намного больше тока во внутрь, чем наружу, в своем рабочем диапазоне напряжений. Эти каналы не являются идеальными выпрямителями, так как они могут пропускать некоторый внешний ток в диапазоне напряжений примерно на 30 мВ выше потенциала покоя.
Эти каналы отличаются от калиевых каналов, которые обычно отвечают за реполяризацию клетки в соответствии с потенциалом действия, например, выпрямителем с задержкой и A-типа. калиевые каналы. Эти более «типичные» калиевые каналы преимущественно проводят наружу (а не внутрь) калиевые токи при деполяризованных мембранных потенциалах, и их можно рассматривать как «внешнее выпрямление». Когда впервые было обнаружено, внутреннее выпрямление было названо «аномальным выпрямлением», чтобы отличать его от выходящих калиевых токов.
Внутренние выпрямители также отличаются от калиевых каналов с тандемными поровыми доменами, которые в значительной степени ответственны за «утечку». «К токи. Некоторые внутренние выпрямители, называемые «слабыми внутренними выпрямителями», переносят измеряемые наружные токи K при напряжениях, положительных по отношению к потенциалу разворота K (соответствующему, но большему, чем небольшие токи над линией 0 нА на рисунке 1). Они вместе с каналами «утечки» устанавливают мембранный потенциал покоя клетки. Другие каналы внутреннего выпрямления, называемые «сильные внутренние выпрямители», вообще несут очень небольшой внешний ток и в основном активны при напряжениях, отрицательных по отношению к потенциалу разворота K, где они несут внутренний ток (гораздо большие токи ниже линии 0 нА на рисунке 1).
Феномен внутренней ректификации каналов K ir является результатом высокоаффинного блока эндогенными полиаминами, а именно спермин, а также ионы магния, которые закупоривают поры канала при положительных потенциалах, что приводит к уменьшению выходных токов. Этот зависящий от напряжения блок полиаминов приводит к эффективной проводимости тока только во внутреннем направлении. Хотя основная идея полиаминового блока понятна, конкретные механизмы остаются спорными.
Для всех K ir каналов требуется фосфатидилинозитол-4,5-бисфосфат (PIP 2) для активации. PIP 2 связывается и непосредственно активирует K ir 2.2 с агонистоподобными свойствами. В этом отношении K ir каналы представляют собой PIP 2ионные каналы, управляемые лигандами.
Kirканалов обнаруживается во многих типах клеток, включая макрофаги, клетки сердца и почки, лейкоциты, нейроны и эндотелиальные клетки. Опосредуя небольшой деполяризующий ток при отрицательных мембранных потенциалах, они помогают установить мембранный потенциал покоя, а в случае группы Kir3 они помогают опосредовать тормозные реакции нейротрансмиттера, но их роли в клеточной физиологии различаются в зависимости от типа клеток:
| Расположение | Функция |
|---|---|
| сердечные миоциты | Kirканалы закрываются при деполяризации, замедляя реполяризацию мембраны и помогая поддерживать более длительный потенциал сердечного действия. Этот тип внутреннего выпрямительного канала отличается от K-каналов выпрямителя с задержкой, которые помогают реполяризовать нервные и мышечные клетки после потенциалов действия ; и каналы утечки калия, которые обеспечивают большую часть основы мембранного потенциала покоя. |
| эндотелиальных клеток | Kirканалов, участвующих в регуляции синтазы оксида азота. |
| почек | Kirэкспортировать излишки калия в собирательные канальцы для удаления с мочой, или, альтернативно, он может участвовать в обратном захвате калия обратно в организм. |
| нейроны и в клетках сердца | активированные G-белком IRK (K ir3) - важные регуляторы, модулируемые нейротрансмиттерами. Мутация в канале GIRK2 приводит к мутации мышей-ткачей.Мутантные мыши "Weaver" атаксичны и демонстрируют опосредованную нейровоспалением дегенерацию их дофаминергических нейронов. По сравнению с неатаксическим контролем, мутанты Weaver имеют дефицит моторной координации и изменения в региональном метаболизме мозга. Мыши Weaver были исследованы в лабораториях, заинтересованных в изучении. развитие нервной системы и заболевания в течение более 30 лет. |
| панкреатические бета-клетки | KАТФ каналы (состоящие из субъединиц Kir6.2 и SUR1 ) контроль высвобождение инсулина. |
Зависимость от напряжения может регулироваться внешним K, внутренним Mg, внутренним АТФ и / или G-белками. P-домены каналов IRK демонстрируют ограниченное сходство последовательностей с таковыми из семейства VIC. Внутренние выпрямители играют роль в установке потенциалов клеточной мембраны, а закрытие Использование этих каналов при деполяризации позволяет возникать длительные потенциалы действия с фазой плато. У внутренних выпрямителей отсутствуют внутренние спирали измерения напряжения, которые можно найти во многих каналах семейства VIC. В некоторых случаях, например, Kir1.1a, Kir6.1 и Kir6.2, было предложено прямое взаимодействие с членом суперсемейства ABC для придания уникальных функциональных и регуляторных свойств гетеромерному комплексу, включая чувствительность к АТФ.. Эти АТФ-чувствительные каналы находятся во многих тканях организма. Они делают активность канала зависимой от цитоплазматического соотношения АТФ / АДФ (увеличение АТФ / АДФ закрывает канал). Рецепторы SUR1 и SUR2 сульфонилмочевины человека (spQ09428 и Q15527, соответственно) представляют собой белки ABC, которые регулируют каналы Kir6.1 и Kir6.2 в ответ на АТФ и CFTR (TC # 3.A.1.208.4 ) может регулировать Kir1.1a.
Кристаллическая структура и функция бактериальных членов семейства IRK-C были определены. KirBac1.1, от Burkholderia pseudomallei, имеет длину 333 аминоацильных остатка (aas) с двумя N-концевыми TMS, фланкирующими P-петлю (остатки 1-150), и C-концевую половину белка. гидрофильный. Он переносит одновалентные катионы с селективностью: K ≈ Rb ≈ Cs ≫ Li ≈ Na ≈ NMGM (протонированный N-метил-D-глюкамин ). Активность подавляется Ba, Ca и низким pH.
Существует семь подсемейств K ir каналов, обозначен как K ir 1 - K ir 7. Каждое подсемейство имеет несколько членов (например, K ir 2.1, K ir 2.2, K ir 2.3 и т. Д.), Которые имеют почти идентичные аминокислотные последовательности у известных млекопитающих. виды.
Kirканалы образуются из гомотетрамерных мембранных белков. Каждая из четырех идентичных белковых субъединиц состоит из двух охватывающих мембрану альфа-спиралей (M1 и M2). Гетеротетрамеры могут образовываться между членами одного и того же подсемейства (то есть K ir 2.1 и K ir 2.3), когда каналы чрезмерно экспрессируются.
| Ген | Белок | Псевдонимы | Ассоциированные субъединицы |
|---|---|---|---|
| KCNJ1 | Kir1,1 | ROMK1 | NHERF2 |
| KCNJ2 | Kir2,1 | IRK1 | Kir2.2, K ir 4.1, PSD-95, SAP97, AKAP79 |
| KCNJ12 | Kir2.2 | IRK2 | Kir2.1 и K ir 2.3 для образования гетеромерного канала, вспомогательная субъединица: SAP97, Veli-1, Veli-3, PSD-95 |
| KCNJ4 | Kir2.3 | IRK3 | Kir2.1 и K ir 2.3 для формирования гетеромерного канала, PSD-95, Chapsyn-110 / PSD-93 |
| KCNJ14 | Kir2,4 | IRK4 | Kir2.1 для образования гетеромерного канала |
| KCNJ3 | Kir3.1 | GIRK1, KGA | Kir3.2, K ir 3.4, K ir 3.5, K ir 3.1 сам по себе не работает |
| KCNJ6 | Kir3.2 | GIRK2 | Kir3.1, K ir 3.3, K ir 3,4 для образования гетеромерного канала |
| KCNJ9 | Kir3,3 | GIRK3 | Kir3.1, K ir 3,2 для образования гетеромерного канала |
| KCNJ5 | Kir3,4 | GIRK4 | Kir3,1, K ir 3,2, K ir 3,3 |
| KCNJ10 | Kir4,1 | Kir1,2 | Kir4,2, K ir 5.1 и K ir 2.1 с образованием гетеромерного каналы |
| KCNJ15 | Kir4.2 | Kir1.3 | |
| KCNJ16 | Kir5.1 | BIR 9 | |
| KCNJ8 | Kir6.1 | KATP | SUR2B |
| KCNJ11 | Kir6.2 | KATP | SUR1, SUR2A и SUR2B |
| KCNJ13 | Kir7.1 | Kir1.4 |
Бертил Хилле (2001). Ионные каналы возбудимых мембран, 3-е изд. (Sinauer: Sunderland, MA), стр. 149 –154. ISBN 0-87893-321-2.