
Войти
Относительно статический мембранный потенциал, который обычно называют нулевым значением трансмембранного напряжения.
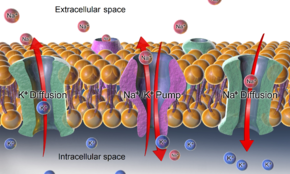 Na + / К + -АТФаза, а также эффекты диффузии вовлеченных ионов, являются основными механизмами поддержания потенциала покоя через мембраны клеток животных.
Na + / К + -АТФаза, а также эффекты диффузии вовлеченных ионов, являются основными механизмами поддержания потенциала покоя через мембраны клеток животных. Относительно статичными мембранный потенциал из покоящихся клеток называется мембранный потенциал покоя (или напряжение покоя), в отличие от конкретных динамических электрохимических явлений, называемых потенциал действия и градуированный мембранный потенциал.
Помимо двух последних, которые встречаются в возбудимых клетках ( нейронах, мышцах и некоторых секреторных клетках в железах ), мембранное напряжение в большинстве невозбудимых клеток также может претерпевать изменения в ответ на внешние или внутриклеточные стимулы. Потенциал покоя существует из - за различий в мембранных проницаемостей для калия, натрия, кальция и хлорид - ионы, которые, в свою очередь, приводит от функциональной активности различных ионных каналов, ионных транспортеров и теплообменников. Обычно мембранный потенциал покоя можно определить как относительно стабильное, основное значение трансмембранного напряжения в клетках животных и растений.
Типичный мембранный потенциал покоя клетки возникает из-за отделения ионов калия от внутриклеточных, относительно неподвижных анионов через мембрану клетки. Поскольку проницаемость мембраны для калия намного выше, чем для других ионов, и из-за сильного химического градиента для калия ионы калия текут из цитозоля во внеклеточное пространство, неся положительный заряд, до тех пор, пока их движение не будет уравновешено накоплением отрицательный заряд на внутренней поверхности мембраны. Опять же, из-за высокой относительной проницаемости для калия результирующий мембранный потенциал почти всегда близок к потенциалу обращения калия. Но для того, чтобы этот процесс произошел, сначала необходимо настроить градиент концентрации ионов калия. Эта работа выполняется ионными насосами / транспортерами и / или теплообменниками и обычно приводится в действие АТФ.
В случае мембранного потенциала покоя через плазматическую мембрану животной клетки градиенты калия (и натрия) устанавливаются Na + / K + -АТФазой (натрий-калиевый насос), которая транспортирует 2 иона калия внутрь и 3 иона натрия снаружи при стоимость 1 молекулы АТФ. В других случаях, например, мембранный потенциал может быть установлен путем подкисления внутренней части мембранного компартмента (такого как протонный насос, который генерирует мембранный потенциал через мембраны синаптических везикул ).
В большинстве количественных обработок мембранного потенциала, таких как вывод уравнения Гольдмана, предполагается электронейтральность ; это означает, что на любой стороне мембраны нет измеримого превышения заряда. Таким образом, хотя существует электрический потенциал на мембране из-за разделения зарядов, нет реально измеримой разницы в общей концентрации положительных и отрицательных ионов на мембране (как это оценивается ниже ), то есть нет реально измеримых превышение заряда с обеих сторон. Это происходит потому, что влияние заряда на электрохимический потенциал намного больше, чем эффект концентрации, поэтому необнаружимое изменение концентрации вызывает большое изменение электрического потенциала.
Клеточные мембраны обычно проницаемы только для части ионов. Обычно они включают ионы калия, ионы хлора, ионы бикарбоната и другие. Чтобы упростить описание ионной основы мембранного потенциала покоя, наиболее полезно сначала рассмотреть только одну разновидность ионов, а позже рассмотреть другие. Поскольку транс-плазматические мембранные потенциалы почти всегда определяются в первую очередь проницаемостью для калия, с этого и следует начать.
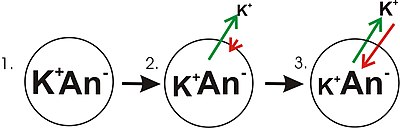 Диаграмма, показывающая прогрессию развития мембранного потенциала от градиента концентрации (для калия). Зеленые стрелки указывают на чистое движение K + вниз по градиенту концентрации. Красные стрелки показывают чистое движение K + из-за мембранного потенциала. Диаграмма вводит в заблуждение в том смысле, что, хотя концентрация ионов калия вне клетки увеличивается, только небольшое количество K + должно пересечь мембрану, чтобы создать мембранный потенциал с величиной, достаточно большой, чтобы противодействовать тенденции ионов калия. для уменьшения градиента концентрации.
Диаграмма, показывающая прогрессию развития мембранного потенциала от градиента концентрации (для калия). Зеленые стрелки указывают на чистое движение K + вниз по градиенту концентрации. Красные стрелки показывают чистое движение K + из-за мембранного потенциала. Диаграмма вводит в заблуждение в том смысле, что, хотя концентрация ионов калия вне клетки увеличивается, только небольшое количество K + должно пересечь мембрану, чтобы создать мембранный потенциал с величиной, достаточно большой, чтобы противодействовать тенденции ионов калия. для уменьшения градиента концентрации. Напряжение покоя является результатом нескольких ферментов, переносящих ионы ( унипортеров, котранспортеров и насосов ) в плазматической мембране, постоянно работающих параллельно, в результате чего каждый ион-транслокатор имеет свою характерную электродвижущую силу (= потенциал реверсирования = 'равновесное напряжение'), в зависимости от конкретных концентраций субстрата внутри и снаружи ( в некоторых насосах включен внутренний АТФ ). H + -экспортирующая АТФаза делает мембранное напряжение в растениях и грибах гораздо более отрицательным, чем в более тщательно изученных клетках животных, где напряжение покоя в основном определяется селективными ионными каналами.
В большинстве нейронов потенциал покоя составляет примерно -70 мВ. Потенциал покоя в основном определяется концентрацией ионов в жидкостях по обе стороны клеточной мембраны и белками переноса ионов, которые находятся в клеточной мембране. Ниже описано, как концентрации ионов и белков мембранного транспорта влияют на значение потенциала покоя.
Потенциал покоя клетки можно наиболее полно понять, рассматривая его в терминах равновесных потенциалов. На приведенной здесь диаграмме в качестве примера модельной ячейке был задан только один проникающий ион (калий). В этом случае потенциал покоя этой клетки будет таким же, как равновесный потенциал для калия.
Однако реальная ячейка более сложна, поскольку она проницаема для многих ионов, каждый из которых вносит свой вклад в потенциал покоя. Чтобы лучше понять, рассмотрим ячейку только с двумя проникающими ионами, калием и натрием. Рассмотрим случай, когда эти два иона имеют одинаковые градиенты концентрации, направленные в противоположных направлениях, и проницаемость мембран для обоих ионов одинакова. K + оставляя клетка будет иметь тенденцию тянуть мембранный потенциал к Е К. Na +, попадающий в клетку, будет стремиться перетянуть мембранный потенциал к потенциалу обращения натрия E Na. Так как в обеих проницаемости ионов были установлены равными, мембранный потенциал будет, в конце Na + / K + перетягивание каната, в конечном итоге на полпути между E Na и E K. Поскольку E Na и E K были равны, но имели противоположные знаки, промежуточное значение равно нулю, что означает, что мембрана будет находиться при 0 мВ.
Обратите внимание, что даже несмотря на то, что мембранный потенциал при 0 мВ является стабильным, это не состояние равновесия, поскольку ни один из участвующих ионов не находится в равновесии. Ионы диффундируют вниз по своим электрохимическим градиентам через ионные каналы, но мембранный потенциал поддерживается постоянным притоком K + и оттоком Na + через переносчики ионов. Такая ситуация с аналогичной проницаемостью для противодействующих ионов, таких как калий и натрий в клетках животных, может быть чрезвычайно дорогостоящей для клетки, если эти проницаемости относительно велики, поскольку для обратной перекачки ионов требуется много энергии АТФ. Поскольку ни одна реальная клетка не может обеспечить такую равную и большую ионную проницаемость в состоянии покоя, потенциал покоя клеток животных определяется преимущественно высокой проницаемостью для калия и доводится до необходимого значения путем регулирования проницаемости и градиентов проницаемости для натрия и хлора.
В здоровой животной клетке проницаемость для Na + составляет около 5% от проницаемости для K + или даже меньше, тогда как соответствующие обратные потенциалы составляют +60 мВ для натрия ( E Na ) и -80 мВ для калия ( E K ). Таким образом, мембранный потенциал не будет точно соответствовать E K, а скорее будет деполяризован от E K на величину примерно 5% от разницы в 140 мВ между E K и E Na. Таким образом, потенциал покоя клетки будет около -73 мВ.
В более формальных обозначениях мембранный потенциал - это средневзвешенное значение равновесного потенциала каждого вносящего вклад иона. Размер каждого груза - это относительная проводимость каждого иона. В нормальном случае, когда три иона вносят вклад в мембранный потенциал:
куда
Для определения мембранных потенциалов два наиболее важных типа мембранных белков-переносчиков ионов - это ионные каналы и переносчики ионов. Белки ионных каналов образуют пути через клеточные мембраны, по которым ионы могут пассивно диффундировать без прямого расхода метаболической энергии. Они обладают селективностью по отношению к определенным ионам, таким образом, существуют селективные по калию, хлоридам и натрию ионные каналы. Различные клетки и даже разные части одной клетки ( дендриты, тела клеток, узлы Ранвье ) будут иметь разное количество различных белков транспорта ионов. Обычно количество определенных калиевых каналов наиболее важно для контроля потенциала покоя (см. Ниже). Некоторые ионные насосы, такие как Na + / K + -ATPase, являются электрогенными, то есть они создают дисбаланс зарядов на клеточной мембране и также могут вносить непосредственный вклад в мембранный потенциал. Большинство помп для работы используют метаболическую энергию (АТФ).
Для большинства клеток животных ионы калия (K +) являются наиболее важными для потенциала покоя. Из-за активного транспорта ионов калия концентрация калия внутри клеток выше, чем снаружи. Большинство клеток имеют белки ионных каналов, селективные по калию, которые остаются открытыми все время. Через эти калиевые каналы будет происходить чистое движение положительно заряженных ионов калия, что приведет к накоплению избыточного отрицательного заряда внутри клетки. Движение положительно заряженных ионов калия наружу происходит из-за случайного молекулярного движения ( диффузии ) и продолжается до тех пор, пока внутри клетки не накопится достаточно избыточного отрицательного заряда, чтобы сформировать мембранный потенциал, который может уравновесить разницу в концентрации калия внутри и снаружи клетки. «Баланс» означает, что электрическая сила ( потенциал ), возникающая в результате накопления ионного заряда и препятствующая внешней диффузии, увеличивается до тех пор, пока не станет равной по величине, но противоположной по направлению тенденции к диффузионному движению калия наружу. Эта точка баланса представляет собой равновесный потенциал, поскольку чистый трансмембранный поток (или ток ) K + равен нулю. Для хорошего приближения равновесного потенциала данного иона необходимы только концентрации по обе стороны от мембраны и температура. Его можно рассчитать с помощью уравнения Нернста :
куда
Равновесные потенциалы калия около -80 милливольт (внутри отрицательные) являются обычными. Различия наблюдаются у разных видов, разных тканей одного и того же животного и одних и тех же тканей в разных условиях окружающей среды. Применяя приведенное выше уравнение Нернста, можно объяснить эти различия изменениями относительной концентрации K + или различиями в температуре.
Для общего использования уравнение Нернста часто приводится в упрощенной форме, принимая типичную температуру человеческого тела (37 ° C), уменьшая константы и переходя на логарифмическую основу 10. (Единицы, используемые для концентрации, не важны, поскольку они сокращаются до соотношение). Для калия при нормальной температуре тела можно рассчитать равновесный потенциал в милливольтах как:
Точно так же потенциал равновесия натрия (Na +) при нормальной температуре человеческого тела рассчитывается с использованием той же упрощенной константы. Вы можете рассчитать E, принимая внешнюю концентрацию [K + ] o, равную 10 мМ, и внутреннюю концентрацию, [K + ] i, равную 100 мМ. Для хлорид-ионов (Cl -) знак константы должен быть изменен на противоположный (−61,54 мВ). При расчете равновесного потенциала для кальция (Ca 2+) заряд 2+ уменьшает упрощенную константу вдвое до 30,77 мВ. При работе при комнатной температуре, около 21 ° C, расчетные константы составляют примерно 58 мВ для K + и Na +, -58 мВ для Cl - и 29 мВ для Ca 2+. При физиологической температуре около 29,5 ° C и физиологических концентрациях (которые различаются для каждого иона) расчетные потенциалы составляют примерно 67 мВ для Na +, -90 мВ для K +, -86 мВ для Cl - и 123 мВ для Ca 2. +.
Мембранный потенциал покоя не является равновесным потенциалом, так как он зависит от постоянного расхода энергии (для ионных насосов, как упоминалось выше) на его поддержание. Этот механизм учитывается динамическим диффузионным потенциалом - полностью в отличие от равновесного потенциала, который справедлив независимо от природы рассматриваемой системы. В мембранном потенциале покоя доминируют ионные частицы в системе, которые имеют наибольшую проводимость через мембрану. Для большинства клеток это калий. Поскольку калий также является ионом с наиболее отрицательным равновесным потенциалом, обычно потенциал покоя может быть не более отрицательным, чем равновесный потенциал калия. Потенциал покоя может быть рассчитан с помощью уравнения напряжения Гольдмана-Ходжкина-Каца с использованием концентраций ионов как для равновесного потенциала, а также с учетом относительной проницаемости каждого вида ионов. В нормальных условиях можно с уверенностью предположить, что только ионы калия, натрия (Na +) и хлорида (Cl -) играют большую роль в потенциале покоя:
Это уравнение напоминает уравнение Нернста, но имеет член для каждого проникающего иона. Кроме того, в уравнение был вставлен z, в результате чего внутриклеточные и внеклеточные концентрации Cl - меняются местами относительно K + и Na +, поскольку отрицательный заряд хлорида обрабатывается путем инвертирования дроби внутри логарифмического члена. * E m - мембранный потенциал, измеренный в вольтах * R, T и F такие, как указано выше * P s - относительная проницаемость иона s * [s] Y - концентрация иона s в отсеке Y, как указано выше. Другой способ увидеть мембранный потенциал, учитывая вместо этого проводимость ионных каналов, а не проницаемость мембраны, - это использовать уравнение Миллмана (также называемое уравнением хордовой проводимости):
или переформулирован
где g tot - общая проводимость всех ионных частиц, снова в произвольных единицах. Последнее уравнение изображает мембранный потенциал покоя как средневзвешенное значение реверсивных потенциалов системы, где веса представляют собой относительные проводимости каждого вида ионов ( г X / г итого ). Во время потенциала действия эти веса изменяются. Если проводимости Na + и Cl - равны нулю, мембранный потенциал уменьшается до потенциала Нернста для K + (как g K + = g tot ). Обычно в условиях покоя g Na + и g Cl− не равны нулю, но они намного меньше, чем g K +, что делает E m близким к E eq, K +. Медицинские условия, такие как гиперкалиемия, в которых в крови в сыворотке калия (который регулирует [K + ] O ) изменяются очень опасны, так как они смещены E э, K +, влияя таким образом на E м. Это может вызвать аритмию и остановку сердца. Использование болюсной инъекции хлорида калия при казнях путем смертельной инъекции останавливает сердце, сдвигая потенциал покоя к более положительному значению, что деполяризует и сокращает сердечные клетки навсегда, не позволяя сердцу реполяризоваться и, таким образом, входить в диастолу для пополнения с кровью.
Хотя уравнение напряжения GHK и уравнение Миллмана связаны, они не эквивалентны. Критическое различие состоит в том, что уравнение Миллмана предполагает, что отношение тока к напряжению является омическим, тогда как уравнение напряжения GHK учитывает небольшие мгновенные выпрямления, предсказываемые уравнением потока GHK, вызванные градиентом концентрации ионов. Таким образом, более точная оценка мембранного потенциала может быть рассчитана с использованием уравнения GHK, чем с помощью уравнения Миллмана.
В некоторых клетках постоянно меняется мембранный потенциал (например, в клетках, задающих ритм сердца ). Для таких клеток никогда не бывает «покоя», а «потенциал покоя» - это теоретическое понятие. Другие клетки с небольшими функциями мембранного транспорта, которые меняются со временем, имеют мембранный потенциал покоя, который можно измерить, вставив электрод в клетку. Трансмембранные потенциалы также можно измерить оптически с помощью красителей, которые изменяют свои оптические свойства в соответствии с мембранным потенциалом.
| Типы клеток | Потенциал отдыха |
|---|---|
| Клетки скелетных мышц | -95 мВ |
| Астроглия | От -80 до -90 мВ |
| Нейроны | От -60 до -70 мВ |
| Гладкомышечные клетки | -60 мВ |
| Гладкая мышечная ткань аорты | -45 мВ |
| Фоторецепторные клетки | -40 мВ |
| Волосковая клетка ( улитка ) | От -15 до -40 мВ |
| Эритроциты | -8,4 мВ |
| Хондроциты | -8 мВ |
Токи покоя в нервах были измерены и описаны Юлиусом Бернштейном в 1902 году, когда он предложил «теорию мембран», которая объяснила потенциал покоя нерва и мышцы как диффузионный потенциал.
 ,
,![E_ {eq, K ^ +} = \ frac {RT} {zF} \ ln \ frac {[K ^ +] _ {o}} {[K ^ +] _ {i}},](https://wikimedia.org/api/rest_v1/media/math/render/svg/b7eb1afcc5d7a5032ff24be5c403b43b1307b01e)
![E _ {{экв, K ^ {+}}} = 61,54 мВ \ log {\ frac {[K ^ {+}] _ {{o}}} {[K ^ {+}] _ {{i}}} },](https://wikimedia.org/api/rest_v1/media/math/render/svg/9d0b62ffda044574b1a52306113028c6a29c89f2)
![E _ {{m}} = {\ frac {RT} {F}} \ ln {\ left ({\ frac {P _ {{Na ^ {+}}} [Na ^ {+}] _ {{o}}) + P _ {{K ^ {+}}} [K ^ {+}] _ {{o}} + P _ {{Cl ^ {-}}} [Cl ^ {-}] _ {{i}}} { P _ {{Na ^ {+}}} [Na ^ {+}] _ {{i}} + P _ {{K ^ {+}}} [K ^ {+}] _ {{i}} + P_ { {Cl ^ {-}}} [Cl ^ {-}] _ {{o}}}} \ right)}](https://wikimedia.org/api/rest_v1/media/math/render/svg/aef443116c02ee0a29c6f4d6f5a5fc571e530ec8)

