
Войти
| формилпептидный рецептор 1 | |
|---|---|
| Идентификаторы | |
| Символ | FPR1 |
| Альт. символы | FPR; FMLP |
| Ген NCBI | 2357 |
| HGNC | 3826 |
| OMIM | 136537 |
| RefSeq | NM_002029 |
| UniProt | P21462 |
| Прочие данные | |
| Locus | Chr. 19 q13.41 |
| рецептор 2 формилпептида | |
|---|---|
| Идентификаторы | |
| Символ | FPR2 |
| Альт. символы | ALXR, FMLPX, FPR2 / ALX, FPR2A, FPRH1, FPRL1, HM63, LXA4R, RFP |
| ген NCBI | 2358 |
| HGNC | 3827 |
| OMIM | 136538 |
| RefSeq | NM_001462 |
| UniProt | P25090 |
| Прочие данные | |
| Locus | Chr. 19 q13.3-13.4 |
| рецептор формилпептида 3 | |
|---|---|
| Идентификаторы | |
| Символ | FPR3 |
| Альт. символы | FPRH2, FPRL2, FMLPY |
| Ген NCBI | 2359 |
| HGNC | 3828 |
| OMIM | 136539 |
| RefSeq | NM_002030 |
| UniProt | P25089 |
| Прочие данные | |
| Locus | Chr. 19 q13.3-13.4 |
рецепторы формилпептида (FPR ) относятся к классу рецепторов, связанных с G-белком участвует в хемотаксисе. У человека существует три изоформы рецептора формилпептида, каждая из которых кодируется отдельным геном, который называется FPR1, FPR2 и FPR3. Эти рецепторы были первоначально идентифицированы по их способности связывать N-формилпептиды, такие как N-формилметионин, продуцируемые разложением либо бактериальных клеток, либо клеток-хозяев. Следовательно, рецепторы формилового пептида участвуют в опосредовании ответа иммунных клеток на инфекцию. Эти рецепторы также могут подавлять иммунную систему при определенных условиях. Тесная филогенетическая связь передачи сигналов в хемотаксисе и обонянии была недавно доказана путем обнаружения белков, подобных рецептору формилпептида, как отдельного семейства хемосенсоров вомероназального органа у мышей.
FPR теперь правильно принято как термин FPR1 Международного союза фундаментальной и клинической фармакологии.
Исследования, проведенные в 1970-х годах, показали, что серия N-формилметионин -содержащих олигопептидов, включая наиболее сильнодействующие и наиболее известные представители этой серии, N-формил-метионил-лейцил-фенилаланин (fMLF или fMet-Leu-Phe), стимулировал нейтрофилы кролика и человека с помощью очевидного рецептор-зависимого механизма, чтобы мигрировать по направленной схеме в классических лабораторных анализах хемотаксиса. Поскольку эти олигопептиды были продуцированы бактериями или синтетическими аналогами таких продуктов, было высказано предположение, что N-формилолигопептиды являются важными хемотатическими факторами, а их рецепторы - важными рецепторами хемотаксического фактора, которые действуют соответственно как сигнальные и сигнализирующие элементы для инициирования воспалительные реакции для защиты от бактериального вторжения. Дальнейшие исследования определили рецептор для N-формилолигопептидов, рецептор формилпептида (FPR), названный так на основании его способности связываться и активироваться под действием олигопептидов. Два рецептора были впоследствии открыты и названы FPR1 и FPR2 на основании сходства предсказанной аминокислотной последовательности их генов с аминокислотной последовательностью FPR, а не на какой-либо способности связываться или активироваться формилолигопептидами. Эти три рецептора были переименованы в FPR1, FPR2 и FPR3, и было обнаружено, что они обладают очень разными специфичностями в отношении формилолигопептидов и очень разными функциями, включая инициирование воспалительных реакций на N-формилпептиды, высвобождаемые не только бактериями, но и множеством высвобождаемых элементов. тканями хозяина; ослабление и разрешение воспалительных реакций; и, возможно, способствуя развитию некоторых неврологических видов рака и ряда неврологических заболеваний Заболевания на основе амилоидов.
Формилпептидный рецептор (FPR) принадлежит к классу рецепторов, обладающих семью гидрофобными трансмембранными доменами. Конформация FPR стабилизируется несколькими взаимодействиями. К ним относятся потенциальное образование солевого мостика между Arg84-Arg205, Lys85-Arg205 и Lys85-Asp284, которые помогают определить трехмерную структуру трансмембранных доменов, а также положительно заряженные остатки (Arg, Lys), которые взаимодействуют с отрицательно заряженными фосфатами. Кроме того, остаток Arg163 может взаимодействовать с лиганд-связывающим карманом второй внеклеточной петли FPR.
Что касается связывания формил-пептида Met-Leu-Phe, существуют дополнительные потенциальные взаимодействия, которые включают взаимодействия водородных связей между Arg84 и Lys85 первой внеклеточной петли и N-формилом группа лиганда, а также пептидный остов формил-Met-Leu-Phe, которые могут образовывать аналогичные взаимодействия. Было показано, что формил-Met фрагмент лиганда образует дисульфидные мостики с остатками Cys, а также продемонстрировано взаимодействие с Arg163. (Важно отметить, что некоторые взаимодействия, которые стабилизируют конформацию рецептора, также могут влиять на связывание лиганда.) Некоторые олигопептиды также были описаны как характерные составляющие, связанные с Asn-ами внеклеточной N-концевой части и с лиганд-связывающим карманом вторая внеклеточная петля. Эти компоненты также могут определять или уточнять взаимодействие лиганд-рецептор.

Схематическая диаграмма рецептора формилпептида 1. Трансмембранные спирали рецептора представлены сине-зеленые цилиндры, тогда как клеточная мембрана, в которую встроен рецептор, изображена желтым. Внеклеточная поверхность клеточной мембраны находится сверху, а внутриклеточная (цитоплазматическая ) поверхность находится снизу. Внеклеточные петли FPR, ответственные за связывание N-for-Met-Leu-Phe (Nfor-MLF), показаны красным.
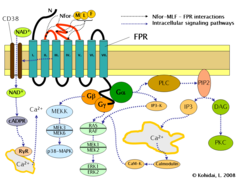
Пути передачи сигналов рецептора формилпептида (FPR).
Индукция FPR запускает множественные изменения в эукариотических клетках, включая перестройку цитоскелета, который в свою очередь, способствует миграции клеток и синтезу хемокинов. Важные пути, регулируемые FPR, включают: