
Войти
| NP_002871. NP_001341618. NP_001341619. NP_001341620. NP_001341600_np1600_np1600_13416001 |
|---|
| Просмотр / редактирование человека | Просмотр / Edit Mouse |
RAF протоонкоген серин / треонин-протеинкиназа, также известный как протоонкоген c-RAF или просто c-Raf или даже Raf-1, представляет собой фермент, который у человека кодируется геном RAF1 . Белок c-Raf является частью пути ERK1 / 2 как MAP-киназа (MAP3K), которая функционирует ниже по течению от подсемейства Ras мембранно-ассоциированных GTPаз. C-Raf является членом семейства киназ Raf из серин / треонин-специфичных протеинкиназ из группы киназ TKL (тирозинкиназоподобных).
Первый Ген Raf, v-Raf был обнаружен в 1983 году. Он был выделен из мышиного ретровируса, имеющего номер 3611. Вскоре было продемонстрировано, что он способен преобразовывать фибробласты грызунов в раковые <25.>клеточные линии, поэтому этот ген получил название быстроускоренной фибросаркомы, индуцированной вирусом (V-RAF). Годом позже в ретровирусе птиц MH2 был обнаружен еще один трансформирующий ген, названный v-Mil, который оказался очень похожим на v-Raf. Исследователи смогли продемонстрировать, что эти гены кодируют ферменты, обладающие активностью серин-треонинкиназы. Нормальные клеточные гомологи v-Raf и v-Mil вскоре были обнаружены как в геноме мыши, так и в геноме курицы (отсюда и название c-Raf для нормального клеточного гена Raf), и стало ясно, что они тоже имели роль в регулировании роста и деления клеток. Теперь мы знаем, что c-Raf является основным компонентом первого описанного пути митоген-активируемой протеинкиназы (MAPK): передача сигналов ERK1 / 2. Он действует как киназа MAP3, инициируя весь каскад киназ. Последующие эксперименты показали, что нормальные клеточные гены Raf также могут мутировать, чтобы стать онкогенами, «перегружая» активность MEK1 / 2 и ERK1 / 2. Фактически, геномы позвоночных содержат несколько генов Raf. Спустя несколько лет после открытия c-Raf были описаны еще две родственные киназы: A-Raf и B-Raf. Последний стал центром исследований в последние годы, поскольку большая часть опухолей человека несут онкогенные «драйверные» мутации в гене B-Raf. Эти мутации вызывают неконтролируемую высокую активность ферментов Raf. Таким образом, диагностический и терапевтический интерес к киназам Raf достиг нового пика в последние годы.
Ген c-Raf человека расположен на хромосоме 3. Описаны по меньшей мере две изоформы мРНК (возникающие в результате включения или удаления альтернативного экзона ), которые демонстрируют лишь незначительные различия. Более короткая основная изоформа, состоящая из 17 экзонов, кодирует протеинкиназу из 648 аминокислот.
 Схематическая архитектура человеческого белка c-Raf
Схематическая архитектура человеческого белка c-Raf Аналогично многим другие MAPKKK, c-Raf представляет собой мультидоменный белок с несколькими дополнительными доменами, способствующими регуляции его каталитической активности. На его N-концевом сегменте Ras-связывающий домен (RBD) и гомологичный домен 1 C-киназы (домен C1) находятся рядом друг с другом. Структуры обоих консервативных доменов были решены в последние десятилетия, что пролило свет на механизмы их регуляции.
Ras-связывающий домен демонстрирует убиквитиноподобную складку (как и многие другие небольшие связывающие G-белки домены) и избирательно связывает только GTP-связанные белки Ras. (Вы можете увидеть это взаимодействие подробно в поле PDB, прикрепленном к статье. Он показывает Rap1 в комплексе с RBD c-Raf.)
Домен C1 - непосредственно ниже по течению Ras-связывающего домена - это особый цинковый палец, богатый цистеинами и стабилизированный двумя ионами цинка. Он подобен диацилглицеринсвязывающим доменам C1 ферментов протеинкиназы C (PKC). Но в отличие от PKC, C1-домены киназ семейства Raf не связывают диацилглицерин. Вместо этого они взаимодействуют с другими липидами, такими как церамид или фосфатидная кислота, и даже способствуют распознаванию активированного Ras (GTP-Ras).
Близость этих двух доменов, а также несколько линий экспериментальных данные свидетельствуют о том, что они действуют как единое целое, отрицательно регулируя активность домена протеинкиназы путем прямого физического взаимодействия. Исторически этот аутоингибиторный блок был обозначен как область CR1 («Консервированная область 1»), шарнирная область была названа CR2, а домен киназы CR3. К сожалению, точная структура аутоингибированной киназы остается неизвестной.
Между блоком аутоингибиторного домена и каталитическим доменом киназы можно найти длинный сегмент, характерный для всех белков Raf. Он сильно обогащен аминокислотами серин, но его точная последовательность плохо консервативна для родственных генов Raf. Эта область кажется изначально неструктурированной и очень гибкой. Его наиболее вероятная роль заключается в том, чтобы действовать как естественный «шарнир» между жестко свернутыми аутоингибиторным и каталитическим доменами, обеспечивая сложные движения и глубокие конформационные перестройки внутри молекулы. Эта шарнирная область содержит небольшой консервативный остров аминокислот, которые отвечают за распознавание 14-3-3 белка, но только когда критический серин (Ser259 в человеческом c-Raf) фосфорилируется. Второй подобный мотив обнаружен на крайнем С-конце (с центром вокруг фосфорилируемого Ser 621) всех ферментов Raf, но ниже киназного домена.
С-концевая половина c-Raf складывается в единый белковый домен, ответственный за каталитическую активность. Структура этого киназного домена хорошо известна как для c-Raf, так и для B-Raf. Он очень похож на другие киназы Raf и белки KSR и отчетливо похож на некоторые другие киназы MAP3, такие как семейство киназ смешанного происхождения (MLK). Вместе они составляют группу протеинкиназ типа тирозинкиназы (TKL). Хотя некоторые особенности объединяют их каталитические домены с протеинтирозинкиназами, активность TKL ограничена фосфорилированием остатков серина и треонина внутри белков-мишеней. Самым важным субстратом киназ Raf (помимо самого себя) являются киназы MKK1 и MKK2, активность которых строго зависит от событий фосфорилирования, выполняемых Rafs.
c-Raf человека является членом более крупного семейства родственных протеинкиназ. Еще два члена, встречающиеся у большинства позвоночных, принадлежат к одному семейству: B-Raf и A-Raf. Помимо разной длины их неконсервативных N- и C-концевых концов, все они имеют одинаковую доменную архитектуру, структуру и регуляцию. По сравнению с относительно хорошо известными c-Raf и B-Raf, очень мало известно о точной функции A-Raf, но также считается, что он аналогичен двум другим членам семейства. Считается, что все эти гены являются продуктом полных дупликаций генов или геномов на заре эволюции позвоночных из одного предкового гена Raf. Большинство других животных организмов обладают только одним геном Raf. Он называется Phl или Draf у дрозофилы и Lin-45 у C. elegans.
 Семейство киназ Raf (схематическая архитектура)
Семейство киназ Raf (схематическая архитектура) Многоклеточные животные также имеют тип киназы, тесно связанный с Raf: это супрессор киназ. Рас (КСР). Позвоночные животные, такие как млекопитающие, имеют два паралогичных гена KSR вместо одного: KSR1 и KSR2. Их C-концевой киназный домен очень похож на Raf (первоначально названный CA5 в KSR и CR3 в Raf), но N-концевой регуляторный регион отличается. Хотя они также имеют перед собой гибкий шарнир (CA4 в KSR) и домен C1 (CA3 в KSR), у KSR полностью отсутствует Ras-связывающий домен. Вместо этого у них есть уникальные регуляторные области на их N-концах, первоначально названные CA1 («консервативная область 1») и CA2. Долгое время структура домена CA1 была загадкой. Однако в 2012 году структура области CA1 в KSR1 была решена: это оказался дивергентный домен SAM (стерильный альфа-мотив), дополненный coiled-coils (CC -SAM): предполагается, что это помогает KSR связываться с мембраной. KSR, подобно Rafs, также имеют двойные связывающие мотивы 14-3-3 (которые зависят от фосфорилирования), но также обладают новыми MAPK-связывающими мотивами на своих шарнирных областях. С типичной последовательностью Phe-x-Phe-Pro (FxFP) эти мотивы важны для регуляции обратной связи киназ Raf в пути ERK1 / 2. Согласно нашим текущим знаниям, KSR также участвуют в том же пути, что и Raf, хотя они играют только вспомогательную роль. Обладая очень низкой собственной киназной активностью, они долгое время считались неактивными, пока их каталитическая активность не была наконец продемонстрирована в последние годы. Но даже тогда они вносят незначительный вклад в фосфорилирование MKK1 и MKK2. Основная роль KSR, по-видимому, заключается в обеспечении партнера для гетеродимеризации ферментов Raf, значительно облегчая их активацию посредством аллостерии. Подобные явления были описаны для других киназ MAP3. ASK2, например, сам по себе является плохим ферментом, и его активность, по-видимому, связана с гетеродимеризацией ASK1 / ASK2.
Raf-подобные киназы полностью отсутствуют в грибах. Но недавнее секвенирование других опистоконтов (например, Capsaspora owczarzaki ) выявило присутствие настоящих киназ Raf у одноклеточных эукариот. Следовательно, возможно, что белки Raf являются древним наследием и предки грибов вторично утратили Raf-зависимую передачу сигналов. Грибковые пути киназ MAP, которые гомологичны пути ERK1 / 2 млекопитающих (Fus3 и Kss1 в дрожжах), активируются связанными с MEKK киназами (например, Ste11 в дрожжах) вместо ферментов Raf.
Raf-киназы, обнаруженные в ретровирусах (таких как мышиный v-Raf), вторично происходят из соответствующих генов позвоночных их хозяев. Эти гены Raf кодируют сильно усеченные белки, в которых отсутствует весь N-концевой аутоингибиторный домен и связывающие мотивы 14-3-3. Известно, что такие серьезные усечения вызывают неконтролируемую активность киназ Raf: это как раз то, что может понадобиться вирусу для эффективного воспроизводства.
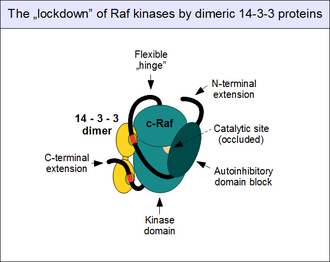 Художественное представление об аутоингибированном состоянии c-Raf, усиленном связанными с ним димерами белка 14-3-3, связанными с фосфорилированными двойными мотивами.
Художественное представление об аутоингибированном состоянии c-Raf, усиленном связанными с ним димерами белка 14-3-3, связанными с фосфорилированными двойными мотивами. Как упоминалось выше, регуляция активности c-Raf сложна. Как «привратник» пути ERK1 / 2, он контролируется множеством тормозных механизмов и обычно не может быть активирован за один шаг. Наиболее важный регуляторный механизм включает прямую физическую ассоциацию N-концевого аутоингибиторного блока с киназным доменом c-Raf. Это приводит к закупорке каталитического сайта и полному отключению киназной активности. Это "закрытое" состояние может быть снято только в том случае, если автоингибирующий домен Raf задействует партнера, конкурирующего с его собственным киназным доменом, наиболее важным из которых является GTP-связанный Ras. Таким образом, активированные малые G-белки могут разрушать внутримолекулярные взаимодействия: это приводит к конформационному изменению («открытию») c-Raf, необходимому для активации киназы и связывания субстрата.
Белки 14-3-3 также вносят вклад в аутоингибирование. Поскольку все белки 14-3-3 образуют конститутивные димеры, их сборки имеют два сайта связывания. Таким образом, димер действует как «молекулярные наручники», запирая их партнеров по связыванию на фиксированном расстоянии и ориентации. Когда точно расположенные двойные связывающие мотивы 14-3-3 задействованы одним димером белка 14-3-3 (например, 14-3-3 дзета), они замыкаются в конформации, которая способствует аутоингибированию и не позволяет разъединению. аутоингибиторного и каталитического доменов. Эта «блокировка» c-Raf (и других Raf, а также KSR) контролируется фосфорилированием мотива. Нефосфорилированные связывающие мотивы 14-3-3 не связываются со своими партнерами: они должны сначала фосфорилироваться по консервативным серинам (Ser 259 и Ser 621) другими протеинкиназами. Наиболее важной киназой, участвующей в этом событии, является активированная TGF-бета киназа 1 (TAK1), а ферментами, предназначенными для удаления этих фосфатов, являются протеинфосфатаза 1 (PP1) и белок комплексы фосфатазы 2A (PP2A).
Обратите внимание, что связывание 14-3-3 ферментов Raf не обязательно является ингибирующим: как только Raf открывается и димеризуется, 14-3-3 также могут связываться в транс, соединяя две киназы и "сковывать наручники" их вместе, чтобы усилить димер, вместо того, чтобы держать их подальше друг от друга. Существуют и другие способы взаимодействия 14-3-3 с c-Raf, но их роль недостаточно хорошо известна.
Димеризация - еще один важный механизм регуляции активности c-Raf, необходимый для петли активации Raf фосфорилирование. Обычно в димеризации участвуют только «открытые» киназные домены. В отличие от B-Raf, который легко образует гомодимеры с самим собой, c-Raf предпочитает гетеродимеризацию либо с B-Raf, либо с KSR1. Гомодимеры и гетеродимеры ведут себя одинаково. Структура домена гомодимерной киназы B-Raf ясно показывает, что петли активации (которые контролируют каталитическую активность всех известных протеинкиназ) расположены в димере в активноподобной конформации. Это связано с аллостерическим эффектом связывания другой молекулы с «обратной» стороной киназы; такие димеры симметричны и имеют два частично активных каталитических центра. На этом этапе активность киназ Raf низкая и нестабильная.
 Цикл активации белков Raf млекопитающих, представленный B-Raf (значительно упрощенный обзор, не показаны все этапы).
Цикл активации белков Raf млекопитающих, представленный B-Raf (значительно упрощенный обзор, не показаны все этапы). Для достижения полной активности и стабилизации активного состояния петля активации c-Raf должна быть фосфорилироваться. Единственные известные в настоящее время киназы, выполняющие этот акт, - это сами киназы семейства Raf. Но некоторые другие киназы, такие как PAK1, могут фосфорилировать другие остатки рядом с киназным доменом c-Raf: точная роль этих вспомогательных киназ неизвестна. В контексте c-Raf и c-Raf, и KSR1 необходимы для стадии «трансфосфорилирования». Из-за архитектуры димеров это фосфорилирование может происходить только в транс (то есть один димер фосфорилирует другой в четырехчленном переходном комплексе). Взаимодействуя с консервативными остатками Arg и Lys в киназном домене, фосфорилированные активационные петли сдвигают конформацию и становятся упорядоченными, постоянно блокируя киназный домен в полностью активном состоянии до тех пор, пока не будет дефосфорилирован. Фосфорилированные петли активации также делают киназу нечувствительной к присутствию ее аутоингибиторного домена. KSR не могут пройти эту последнюю стадию, так как они пропускают какие-либо фосфорилируемые остатки в своих петлях активации. Но как только c-Raf полностью активирован, в этом больше нет необходимости: активные ферменты Raf теперь могут взаимодействовать со своими субстратами. Как и большинство протеинкиназ, c-Raf имеет несколько субстратов. BAD (Bcl2-атагонист клеточной гибели) непосредственно фосфорилируется c-Raf вместе с несколькими типами аденилатциклазы, миозинфосфатазы (MYPT), тропонин Т сердечной мышцы (TnTc) и т.д. В качестве возможных субстратов также были предложены белок ретинобластомы (pRb) и фосфатаза Cdc25.
Наиболее важными мишенями для всех ферментов Raf являются MKK1 (MEK1) и MKK2 (MEK2). Хотя структура фермент-субстратного комплекса c-Raf: MKK1 неизвестна, ее можно точно смоделировать по образцу комплекса KSR2: MKK1. Здесь не происходит реального катализа, но считается, что он очень похож на способ связывания Raf со своими субстратами. Главный интерфейс взаимодействия обеспечивается С-концевыми долями обоих киназных доменов; большая, неупорядоченная, богатая пролином петля, уникальная для MKK1 и MKK2, также играет важную роль в ее позиционировании по отношению к Raf (и KSR). Эти MKK фосфорилируются по крайней мере на двух сайтах в их петлях активации после связывания с Raf: это также активирует их. Мишенями киназного каскада являются ERK1 и ERK2, которые избирательно активируются MKK1 или MKK2. ERK имеют множество субстратов в клетках; они также способны перемещаться в ядро для активации ядерных факторов транскрипции. Активированные ERK являются плейотропными эффекторами клеточной физиологии и играют важную роль в контроле экспрессии генов, участвующих в цикле клеточного деления, миграции клеток, ингибировании апоптоза и дифференцировке клеток.
Наследственные мутации c-Raf, связанные с повышением функции, участвуют в некоторых редких, но тяжелых синдромах. Большинство этих мутаций связано с заменой одной аминокислоты в одном из двух связывающих мотивов 14-3-3. Мутация c-Raf является одной из возможных причин синдрома Нунана : у пораженных людей врожденные пороки сердца, низкий и дисморфический рост и несколько других деформаций. Подобные мутации в c-Raf могут также вызывать родственное состояние, называемое синдромом LEOPARD (лентиго, электрокардиографические аномалии, глазной гипертелоризм, легочный стеноз, аномальные гениталии, задержка роста, глухота) со сложной ассоциацией дефектов.
Хотя c-Raf явно способен к мутации в онкоген в экспериментальных условиях и даже в некоторых опухолях человека, его сестринская киназа B-Raf является истинным основным
Примерно 20% всех исследованных образцов опухолей человека содержат мутированный ген B-Raf. Подавляющее большинство этих мутаций связано с заменой одной аминокислоты: Val 600 на Glu, и этот аберрантный генный продукт (BRAF-V600E) можно визуализировать с помощью иммуногистохимии для клинической молекулярной диагностики. Аберрация может имитировать фосфорилирование петли активации и - с помощью переход через все шаги управления при нормальной активации - немедленно сделать киназный домен полностью активным. Поскольку B-Raf может также активироваться посредством гомодимеризации, а c-Raf - посредством гетеродимеризации, эта мутация имеет катастрофический эффект, превращая путь ERK1 / 2 в конститутивно активный и управляя неконтролируемым процессом деления клеток.
Из-за важности мутаций Ras и B-Raf в онкогенезе были разработаны несколько ингибиторов Raf для борьбы с раком, особенно против B-Raf, проявляющего мутацию V600E. Сорафениб был первым клинически полезным агентом, который представляет собой фармакологическую альтернативу для лечения ранее неизлечимых злокачественных новообразований, таких как почечно-клеточная карцинома и меланома. Следили за несколькими другими молекулами, такими как вемурафениб, регорафениб, дабрафениб и т.д.
К сожалению, АТФ-конкурентный B- Ингибиторы Raf могут иметь нежелательный эффект при K-Ras-зависимом раке: они просто слишком селективны для B-Raf. Хотя они отлично ингибируют активность B-Raf в случае, если мутантный B-Raf является основным виновником, они также способствуют гомо- и гетеродимеризации B-Raf с самим собой и c-Raf. Это фактически усилит активацию c-Raf вместо ее ингибирования в случае, если нет мутации в каких-либо генах Raf, но их общий активаторный белок K-Ras является мутированным. Эта «парадоксальная» активация c-Raf делает необходимым проведение скрининга мутаций B-Raf у пациентов (с помощью генетической диагностики) перед началом терапии ингибитором B-Raf.
Было показано, что C-Raf взаимодействует с:



