
Войти
Каналродопсины являются подсемейством ретинилиденовых белков (родопсинов ), которые функционируют как светозависимые ионные каналы. Они служат в качестве сенсорных фоторецепторов в одноклеточных зеленых водорослях, контролируя фототаксис : движение в ответ на свет. Экспрессируемые в клетках других организмов, они позволяют свету контролировать электрическую возбудимость, внутриклеточную кислотность, приток кальция и другие клеточные процессы ( см. оптогенетика ). Каналродопсин-1 (ChR1) и каналродопсин-2 (ChR2) из модельного организма Chlamydomonas reinhardtii являются первыми обнаруженными канальными родопсинами. Были клонированы варианты из других видов водорослей, и ожидается их появление.
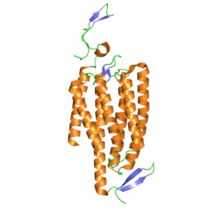 Кристаллическая структура канального родопсина. PDB 3ug9
Кристаллическая структура канального родопсина. PDB 3ug9 С точки зрения структуры канальные родопсины представляют собой ретинилиденовые белки. Они представляют собой семистрансмембранные белки, подобные родопсину, и содержат изомеризуемый светом хромофор все- транс - ретиналь (альдегид производное витамина A ). Хромофор сетчатки ковалентно связан с остальным белком через протонированное основание Шиффа. В то время как большинство 7-трансмембранных белков являются рецепторами, связанными с G-белками, которые открывают другие ионные каналы косвенно через вторичные мессенджеры (т.е. они метаботропны ), канальные родопсины непосредственно образуют ионные каналы (т.е. они ионотропны ). Это делает клеточную деполяризацию чрезвычайно быстрой, надежной и полезной для приложений биоинженерии и нейробиологии, включая фотостимуляцию.
 Схема слитой конструкции ChR2-RFP
Схема слитой конструкции ChR2-RFP Природный («дикого типа») ChR2 поглощает синий свет с максимумом спектра поглощения и действия при 480 нм. Когда полностью транс-ретинальный комплекс поглощает фотон, он вызывает конформационные изменения от полностью транс-ретиналя к 13-цис-ретиналю. Это изменение вводит еще один в трансмембранный белок, открывая поры по крайней мере до 6 Å. В течение миллисекунд сетчатка возвращается в полностью трансформированную форму, закрывая поры и останавливая поток ионов. Большинство естественных канальных родопсинов представляют собой неспецифические катионные каналы, проводящие ионы H, Na, K и Ca. Недавно были открыты анион-проводящие родопсины.
Канальные родопсины являются ключевыми инструментами в оптогенетике. С-концевой конец канала родопсина-2 простирается во внутриклеточное пространство и может быть заменен флуоресцентными белками, не влияя на функцию канала. Этот вид слитой конструкции может быть полезен для визуализации морфологии клеток, экспрессирующих ChR2. Было показано, что точечные мутации вблизи кармана связывания сетчатки влияют на биофизические свойства канального родопсина, в результате чего в различных инструментах.
Закрытие канала после оптической активации можно существенно отсрочить путем мутации белковых остатков C128 или D156. Эта модификация приводит к получению сверхчувствительных каналов родопсинов, которые могут открываться синим световым импульсом и закрываться зеленым или желтым световым импульсом (ступенчатые опсины). Мутация остатка E123 ускоряет кинетику канала (ChETA), и полученные в результате мутанты ChR2 были использованы для спайк-нейронов с частотой до 200 Гц. В целом, родопсины с медленной кинетикой более светочувствительны на уровне популяции, поскольку открытые каналы со временем накапливаются даже при слабом освещении.
Мутанты H134R и T159C демонстрируют повышенные фототоки, а комбинация T159 и E123 (ET / TC) имеет немного большие фототоки и немного более быструю кинетику, чем ChR2 дикого типа. Среди вариантов ChR, ChIEF, химера и точечный мутант ChR1 и ChR2, демонстрирует наибольшие фототоки и наименьшую десенсибилизацию и имеет кинетику, аналогичную ChR2 дикого типа.
Химерные канальные родопсины имеют были разработаны путем объединения трансмембранных спиралей из ChR1 и VChR1, что привело к развитию ChR с красными спектральными сдвигами (таких как C1V1 и ReaChR). ReaChR улучшает мембранный перенос и сильную экспрессию в клетках млекопитающих и используется для минимально инвазивной транскраниальной активации ствола мозга мотонейронов. Поиски гомологичных последовательностей у других организмов привели к получению спектрально улучшенных и более сильных канальных родпсинов с красным смещением (Chrimson). В сочетании с ChR2 эти желто-красные светочувствительные канальные родопсины позволяют независимо контролировать две популяции нейронов с помощью световых импульсов разного цвета.
Каналродопсин со смещенным синим сдвигом был обнаружен в водоросли Scherffelia dubia. После некоторых разработок, направленных на улучшение движения и скорости мембран, полученный инструмент (CheRiff) произвел большие фототоки при возбуждении 460 нм. Он был объединен с генетически закодированным индикатором кальция jRCaMP1b в полностью оптической системе под названием OptoCaMP.
Мутация L132C (CatCh) увеличивает проницаемость для кальция и генерирует очень большие токи. Мутация E90 в положительно заряженную аминокислоту аргинин превращает канал родопсин из неспецифического катионного канала в хлорид-проводящий канал (ChloC). Селективность по Cl- была дополнительно улучшена за счет замены отрицательно заряженных остатков в поре канала, что сделало реверсивный потенциал более отрицательным. Селективные хлорид-проводящие каналы родопсинов (iChloC, iC ++, GtACR) подавляют выброс нейронов в культуре клеток и у интактных животных при освещении синим светом.
Каналродопсины могут быть легко экспрессированы в возбудимых клетках, таких как нейроны, с использованием различных методов трансфекции (вирусная трансфекция, электропорация, генная пушка ) или трансгенных животных. Поглощающий свет пигмент сетчатки присутствует в большинстве клеток (позвоночных ) в виде витамина А, что позволяет фотостимулировать нейроны без добавление любых химических соединений. До открытия канальных родопсинов нейробиологи ограничивались записью активности нейронов в головном мозге и коррелированием этой активности с поведением. Этого недостаточно, чтобы доказать, что записанная нейронная активность действительно вызвала такое поведение. Управление сетями генетически модифицированных клеток с помощью света, новая область, известная как оптогенетика., Позволяет исследователям теперь исследовать причинную связь между активностью в определенной группе нейронов и психическим события, например принятие решений. Оптический контроль поведения был продемонстрирован у нематод, плодовых мух, рыбок данио и мышей. Недавно были разработаны хлорпроводящие канальные родопсины, которые также были обнаружены в природе. Эти инструменты могут быть использованы для подавления нейронов в культуре клеток и у живых животных с помощью подавления шунтирования.
Использование нескольких цветов света расширяет возможности оптогенетических экспериментов. ChR2, чувствительный к синему свету, и активируемый желтым светом хлоридный насос галлородопсин вместе обеспечивают многоцветную оптическую активацию и подавление нейронной активности. VChR1 из колониальной водоросли Volvox carteri поглощает максимально при 535 нм и использовался для стимуляции клеток желтым светом (580 нм), хотя фототоки, генерируемые VChR1, обычно очень малы. Однако гибриды VChR1-ChR2 были разработаны с использованием направленной эволюции, которые демонстрируют максимальное возбуждение при 560 нм и 50% пикового поглощения при длинах волн более 600 нм.
Используя флуоресцентно меченый ChR2, свет -стимулированные аксоны и синапсы могут быть идентифицированы. Это полезно для изучения молекулярных событий во время индукции синаптической пластичности. Трансфицированные культивируемые нейронные сети можно стимулировать для выполнения некоторых желаемых действий для приложений в робототехнике и управлении. ChR2 также использовался для сопоставления дальнодействующих соединений от одной стороны мозга к другой и для сопоставления пространственного расположения входов на дендритном дереве отдельных нейронов.
Зрительная функция у слепых мышей может быть частично восстанавливается путем экспрессии ChR2 во внутренних клетках сетчатки. В будущем ChR2 может найти применение в медицине, например при формах дегенерации сетчатки или для глубокой стимуляции мозга. Оптические кохлеарные имплантаты показали свою эффективность в экспериментах на животных и могут привести к первому применению оптогенетики у людей.
Подвижность и фотоориентация микроводорослей (фототаксис ) изучались более ста лет во многих лабораториях по всему миру.
В 1980 году Кен Фостер разработал первую последовательную теорию о функциональности глаз водорослей. Он также проанализировал опубликованные спектры действия и дополнил слепые клетки сетчаткой и аналогами сетчатки, что привело к заключению, что фоторецептор для реакции подвижности у Chlorophyceae - это родопсин.
Фототоки Chlorophyceae Heamatococcus pluvialis и Chlamydomonas reinhardtii изучались в течение многих лет в группах Олега Синещекова и Петера Хегеманна, в результате чего в 1978 и 1991 годах были опубликованы две основополагающие публикации. На основе спектроскопии действия и одновременной регистрации фототоков и биения жгутиков было установлено определили, что токи фоторецепторов и последующие движения жгутиков опосредуются родопсином и контролируют фототаксис и фотофобные реакции. Чрезвычайно быстрое нарастание тока фоторецептора после короткой вспышки света привело к выводу, что родопсин и канал тесно связаны в белковый комплекс или даже внутри одного белка.
Однако биохимическая очистка родопсин-фоторецепторы были безуспешными в течение многих лет.
Нуклеотидные последовательности родопсинов, которые теперь называются канальными родопсинами ChR1 и ChR2, были наконец обнаружены в крупномасштабном проекте секвенирования EST в C. reinhardtii. Независимое представление одних и тех же последовательностей в GenBank тремя исследовательскими группами вызвало путаницу в их именовании: имена cop-3 и cop-4 использовались для первоначального представления группой Хегеманна; csoA и csoB группы Спудича; и acop-1 и acop-2 группы Такахаши. Георгом Нагелем, Эрнстом Бамбергом, Питером Хегеманном и другими было обнаружено, что обе последовательности функционируют как однокомпонентные светоактивированные катионные каналы в ооцитах Xenopus и клетках почек человека (HEK).
Название «каналродопсин» было были придуманы, чтобы подчеркнуть это необычное свойство, и последовательности были соответственно переименованы. Между тем их роль в генерации фоторецепторных токов в клетках водорослей охарактеризовали Олег Синещеков, Кван-Хван Юнг и Джон Спудич, а также Питер Бертольд и Питер Хегеманн.
В ноябре 2004 г. Чжуо-Хуа Пань представил в Nature статью о восстановлении зрения у слепых мышей, трансфицированных каналомродопсином, но статья была отклонена и в конечном итоге опубликована в Neuron в 2006 году.
Между тем, в 2005 году три группы последовательно установили ChR2 в качестве инструмента для генетически целевого оптического дистанционного управления (оптогенетика ) нейронами, нейронными цепями и поведением.
Сначала лаборатория Карла Дейссерота (в статье, опубликованной в августе 2005 г.) продемонстрировала, что ChR2 можно использовать для управления нейронами млекопитающих in vitro., достигая временной точности порядка миллисекунд (как с точки зрения задержки пиков, так и с точки зрения временного дрожания). Это было важным открытием, поскольку, во-первых, для всех опсинов (микробных и позвоночных) требуется сетчатка в качестве светочувствительного кофактора, и было неясно, будут ли центральные нервные клетки млекопитающих содержать достаточные уровни сетчатки. но они делают; во-вторых, он показал, несмотря на небольшую одноканальную проводимость, достаточную эффективность, чтобы управлять нейронами млекопитающих выше порога потенциала действия; и, в-третьих, он продемонстрировал, что каналродопсин является первым оптогенетическим инструментом, с помощью которого нейронная активность может контролироваться с временной точностью, с которой работают нейроны (миллисекунды). Более ранний инструмент для фотостимуляции, cHARGe, продемонстрировал доказательство принципа действия на культивируемых нейронах, но никогда не использовался другими группами, так как он работал с точностью порядка секунд, сильно варьировал и не позволял контролировать индивидуальные потенциалы действия..
Второе исследование было опубликовано позже группами Питера Хегеманна и Стефана Херлитце, подтвердившего способность ChR2 контролировать активность нейронов позвоночных в это время в спинном мозге цыпленка. Это исследование было первым, в котором ChR2 экспрессировался вместе с оптическим глушителем, в данном случае позвоночным родопсином -4, впервые продемонстрировав, что возбудимые клетки могут быть активированы и заглушены с использованием этих двух инструментов одновременно, освещая ткань. на разных длинах волн.
Группы Александра Готтшалка и Эрнста Бамберга (во главе с Георгом Нагелем) продемонстрировали, что ChR2, если он экспрессируется в определенных нейронах или мышечных клетках, может вызывать предсказуемое поведение, то есть может контролировать нервную систему неповрежденного человека. животное, в данном случае беспозвоночное C. elegans. Это было первое использование ChR2 для управления поведением животного в оптогенетическом эксперименте, при котором генетически определенный тип клеток подвергался оптическому дистанционному контролю. Хотя оба аспекта были проиллюстрированы ранее в том же году другой группой, лабораторией Miesenböck, использовавшей косвенно управляемый светом ионный канал P2X2, отныне именно микробные опсины, такие как каналродопсин, доминировали в области генетически ориентированного дистанционного контроля возбудимых клеток, благодаря мощности, скорости, целенаправленности, простоте использования и временной точности прямой оптической активации, не требующей каких-либо внешних химических соединений, таких как лиганды в клетках.
Чтобы преодолеть его основные недостатки - небольшой одиночный -канальная проводимость (особенно в установившемся режиме), ограничение одной оптимальной длиной волны возбуждения (~ 470 нм, синий цвет), а также относительно длительное время восстановления, не позволяющее управлять возбуждением нейронов выше 20-40 Гц - ChR2 был оптимизирован с использованием генной инженерии. точечная мутация H134R (замена аминокислоты гистидина в положении 134 нативного белка на аргинин) привела к увеличению стационарной проводимости, как описано в статье 2005 года, в которой также установлено, что ChR2 является оптогенетическим инструментом в C. elegans. В 2009 году лаборатория Роджера Цзяня оптимизировала ChR2 для дальнейшего увеличения стабильной проводимости и резко снизила десенсибилизацию путем создания химер ChR1 и ChR2 и мутации определенных аминокислот, в результате чего были получены ChEF и ChIEF, которые позволили управлять цепей потенциалов действия до 100 Гц. В 2010 году группы Hegemann и Deisseroth ввели мутацию E123T в нативный ChR2, в результате чего появился ChETA, который имеет более быструю кинетику включения и выключения , позволяя контролировать индивидуальные потенциалы действия на частотах до 200 Гц ( в соответствующих типах клеток).
Группы Hegemann и Deisseroth также обнаружили, что введение точечной мутации C128S превращает полученную ChR2-производную в инструмент ступенчатой функции: после "включения" синим светом ChR2 (C128S) остается в открытом состоянии до тех пор, пока не будет выключен желтым светом - модификация, которая ухудшает временную точность, но увеличивает светочувствительность на два порядка. Они также обнаружили и охарактеризовали ВЧР1 в многоклеточных водорослях Volvox carteri. VChR1 производит только крошечные фототоки, но со спектром поглощения, который смещен в красную сторону относительно ChR2. Используя части последовательности ChR1, амплитуда фототока была позже улучшена, чтобы обеспечить возбуждение двух популяций нейронов на двух разных длинах волн.
Группа Деиссерота впервые применила множество приложений для живых животных, таких как генетически направленное дистанционное управление у грызунов. in vivo, оптогенетическая индукция обучения у грызунов, экспериментальное лечение болезни Паркинсона у крыс и комбинация с фМРТ (опто-фМРТ). Другие лаборатории первыми использовали комбинацию стимуляции ChR2 с визуализацией кальция для полностью оптических экспериментов, картирования дальнодействующих и локальных нервных цепей, экспрессии ChR2 из трансгенного локуса - напрямую или в условной парадигме Cre-lox - а также двухфотонное возбуждение ChR2, позволяющее активировать отдельные клетки.
В марте 2013 года премия мозга (Европейская премия Грете Лундбек по исследованию мозга) была присуждена совместно Бамбергу, Бойдену, Дейссероту, Хегеману, Мизенбёку и Нагелю за «их изобретение и усовершенствование оптогенетики». В том же году Хегеманн и Нагель получили Премию Луи-Жанте в области медицины за «открытие канального родопсина». В 2015 году Бойден и Дайссерот получили премию за прорыв в науках о жизни, а в 2020 году Мизенбёк, Хегеманн и Нагель получили премию Шоу в области наук о жизни и медицине за развитие оптогенетики.