
Войти
| Альфа-мотонейрон | |
|---|---|
 Альфа-мотонейрон происходит из базальной пластинки (базальной пластинки) развивающийся эмбрион. Альфа-мотонейрон происходит из базальной пластинки (базальной пластинки) развивающийся эмбрион. | |
| Идентификаторы | |
| NeuroLex ID | sao1154704263 |
| TH | H2.00.01.0.00008 |
| FMA | 83664 |
| Анатомические термины нейроанатомии [редактировать в Викиданных ] | |
Альфа (α) моторные нейроны (также называемые альфа-мотонейронами ), большие, многополярные нижние моторные нейроны ствола мозга и спинной мозг. Они иннервируют экстрафузальные мышечные волокна скелетных мышц и непосредственно ответственны за начало их сокращения. Альфа-мотонейроны отличаются от гамма-мотонейронов, которые иннервируют интрафузальные мышечные волокна мышечных веретен.
, тогда как их клеточные тела находятся в центральная нервная система (ЦНС), α мотонейроны также считаются частью соматической нервной системы - ветви периферической нервной системы (ПНС) - потому что их аксоны проходят на периферию, чтобы иннервировать скелетные мышцы.
Альфа-мотонейрон и мышечные волокна, которые он иннервирует, являются моторной единицей. Пул моторных нейронов содержит тела всех альфа-мотонейронов, участвующих в сокращении одной мышцы.
Альфа-мотонейроны (α-МН), иннервирующие голову и шея находятся в стволе мозга ; остальные α-MN иннервируют остальную часть тела и находятся в спинном мозге. В спинном мозге больше α-МН, чем в стволе мозга, поскольку количество α-МН прямо пропорционально степени контроля мелкой моторики в этой мышце. Например, мышцы одного пальца имеют больше α-MN на волокно и больше α-MN в целом, чем мышцы четырехглавой мышцы, что позволяет более точно контролировать силу, прилагаемую пальцем.
Как правило, α-МН на одной стороне ствола или спинного мозга иннервируют мышцы на той же стороне тела. Исключение составляет блокированное ядро в стволе мозга, которое иннервирует верхнюю косую мышцу глаза на противоположной стороне лица.
В стволе мозга α-MN и другие нейроны находятся в кластерах клеток, называемых ядрами, некоторые из которых содержат тела клеток. нейронов черепных нервов. Не все ядра черепных нервов содержат α-MN; те, что есть, являются двигательными ядрами, а другие - сенсорными ядрами. Моторные ядра находятся по всему стволу мозга - мозговому веществу, мосту и среднему мозгу - и по причинам развития находятся около средней линии ствола мозга.
Как правило, двигательные ядра, расположенные выше в стволе мозга (т.е. более ростральные), иннервируют мышцы, расположенные выше на лице. Например, глазодвигательное ядро содержит α-МН, которые иннервируют мышцы глаза, и находится в среднем мозге, самом ростральном компоненте ствола мозга. Напротив, подъязычное ядро , которое содержит α-МН, иннервирующие язык, находится в продолговатом мозге, наиболее каудальной (то есть в нижней части) структур ствола мозга.
 Кортикоспинальный тракт является одним из основных нисходящих путей от головного мозга к α-МН спинного мозга.
Кортикоспинальный тракт является одним из основных нисходящих путей от головного мозга к α-МН спинного мозга.В спинном мозге α-МН расположены в сером веществе, которое образует вентральный рог. Эти α-МН обеспечивают двигательный компонент спинномозговых нервов, которые иннервируют мышцы тела.
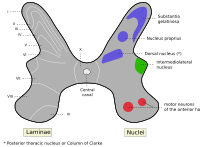 Альфа-мотонейроны расположены в ламине IX в соответствии с системой пластинок Рекседа..
Альфа-мотонейроны расположены в ламине IX в соответствии с системой пластинок Рекседа..Как и в стволе мозга, более высокие сегменты спинного мозга содержат альфа-МН, которые иннервируют мышцы, расположенные выше на теле. Например, двуглавая мышца плеча, мышца руки, иннервируется α-МН в сегментах C5, C6 и C7 спинного мозга, которые находятся рострально в спинном мозге. С другой стороны, икроножная мышца, одна из мышц ноги, иннервируется α-МН в сегментах S1 и S2, которые находятся каудально в спинном мозге.
Альфа-мотонейроны расположены в определенной области серого вещества спинного мозга. Эта область обозначена lamina IX в системе пластинок Rexed, которая классифицирует области серого вещества на основе их цитоархитектуры. Lamina IX располагается преимущественно в медиальной части вентрального рога, хотя есть некоторый вклад в lamina IX за счет набора двигательных нейронов, расположенных более латерально. Как и другие области спинного мозга, клетки в этой пластинке соматотопически организованы, что означает, что положение нейронов в спинном мозге связано с тем, какие мышцы они иннервируют. В частности, α-МН в медиальной зоне ламины IX имеют тенденцию иннервировать проксимальные мышцы тела, в то время как те, что в боковой зоне, имеют тенденцию иннервировать более дистальные мышцы. Аналогичная соматотопия связана с α-МН, которые иннервируют мышцы-сгибатели и разгибатели: α-МН, которые иннервируют сгибатели, как правило, располагаются в дорсальной части пластинки IX; те, что иннервируют разгибатели, как правило, расположены более вентрально.
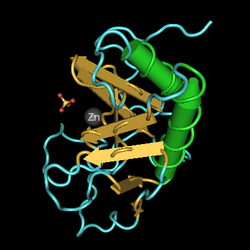 Под воздействием белка sonic hedgehog, показанного здесь, клетки пластинки дна развивающегося спинного мозга дифференцируются в альфа-мотонейроны.
Под воздействием белка sonic hedgehog, показанного здесь, клетки пластинки дна развивающегося спинного мозга дифференцируются в альфа-мотонейроны. Альфа-мотонейроны берут начало в базальной пластинке, вентральной части нервной трубки развивающегося эмбриона. Sonic hedgehog (Shh) секретируется ближайшей хордой и другими вентральными структурами (например, пластиной дна ), создавая градиент высокой концентрации Shh в базальная пластинка и менее концентрированная Shh в крыловой пластине. Под влиянием Shh и других факторов некоторые нейроны базальной пластинки дифференцируются в α-MN.
Как и другие нейроны, α-МН посылают аксональные проекции для достижения своей цели экстрафузальных мышечных волокон через наведение аксонов, процесс частично регулируется нейротрофическими факторами, высвобождаемыми целевыми мышечными волокнами. Нейротрофические факторы также гарантируют, что каждое мышечное волокно иннервируется соответствующим количеством α-МН. Как и большинство типов нейронов в нервной системе, α-MN более многочисленны на раннем этапе развития, чем во взрослом возрасте. Мышечные волокна секретируют ограниченное количество нейротрофических факторов, способных поддерживать только часть α-МН, которые первоначально проецируются в мышечные волокна. Те α-МН, которые не получают достаточного количества нейротрофических факторов, будут подвергаться апоптозу, форме запрограммированной гибели клеток.
Поскольку они иннервируют многие мышцы, некоторые кластеры α-МН получают высокие концентрации нейротрофических факторы и выживают на этой стадии отсечения нейронов. Это верно для α-МН, иннервирующих верхние и нижние конечности: эти α-МН образуют большие столбцы клеток, которые способствуют шейному и поясничному увеличению спинного мозга. Помимо получения нейротрофических факторов от мышц, α-МН также секретируют ряд трофических факторов для поддержки мышечных волокон, которые они иннервируют. Пониженный уровень трофических факторов способствует атрофии мышц, которая следует за поражением α-МН.
Как и другие нейроны, нижние двигательные нейроны имеют как афферентные (входящие), так и эфферентные (исходящие) связи. Альфа-мотонейроны получают входные данные от ряда источников, включая верхние мотонейроны, сенсорные нейроны и интернейроны. Первичный выход α-МН - в экстрафузальные мышечные волокна. Эта афферентная и эфферентная связь необходима для достижения скоординированной мышечной активности.
| происхождение UMN | цель α-MN | Название тракта |
|---|---|---|
| Кора головного мозга | Ствол мозга | Кортикоядерный тракт |
| Кора головного мозга | Спинной мозг | Кортикоспинальный тракт |
| Красное ядро | Спинной мозг | Руброспинальный тракт |
| Вестибулярные ядра | Спинной мозг | Вестибулоспинальный тракт |
| Тектум среднего мозга | Спинной мозг | Тектоспинальный тракт |
| Ретикулярная формация | Спинной мозг | Ретикулоспинальный тракт |
Верхние двигательные нейроны (UMNs) посылают входные данные в α-MN несколькими путями, включая (но не ограничиваясь) кортикоядер, кортикоспинальный и руброспинальный тракт. Кортикоядерный и кортикоспинальный тракты обычно встречаются при исследованиях взаимодействия верхних и нижних мотонейронов при контроле произвольных движений.
кортиконуклеарный тракт назван так потому, что он соединяет кору головного мозга с ядрами черепных нервов. (Кортиконуклеарный тракт также называется кортикобульбарным трактом, поскольку мишень в стволе мозга - продолговатый мозг - архаично называется «луковицей».) Именно по этому пути верхние мотонейроны спускаются из коры. и синапс на α-МН ствола мозга. Точно так же UMN коры головного мозга напрямую контролируют α-MN спинного мозга через латеральный и вентральный кортикоспинальный тракты.
Сенсорный вход в α- MNs обширны и происходят из органов сухожилия Гольджи, мышечных веретен, механорецепторов, терморецепторов и других сенсорных нейронов. на периферии. Эти связи обеспечивают структуру нейронных цепей, лежащих в основе рефлексов. Существует несколько типов рефлекторных цепей, самый простой из которых состоит из одного синапса между сенсорным нейроном и α-МН. рефлекс коленного рефлекса является примером такого моносинаптического рефлекса.
Самый обширный вход в α-МН поступает от локальных интернейронов, которые являются наиболее многочисленным типом нейронов в спинном мозге. Среди их многих функций - синапсы интернейронов на α-MN для создания более сложных рефлекторных схем. Одним из типов интернейронов является клетка Реншоу.
Альфа-мотонейроны посылают волокна, которые в основном синапсы на экстрафузальные мышечные волокна. Другие волокна синапса α-MNs на клетках Реншоу, то есть тормозящие интернейроны, которые синапсируют на α-MN и ограничивают его активность, чтобы предотвратить повреждение мышц.
Как и другие нейроны, α-MN передают сигналы в виде потенциалов действия, быстрых изменений электрической активности, которые распространяются от тела клетки к конец аксона . Чтобы увеличить скорость перемещения потенциалов действия, аксоны α-MN имеют большой диаметр и сильно миелинизированы как олигодендроцитами, так и шванновскими клетками. Олигодендроциты миелинизируют часть аксона α-MN, которая находится в центральной нервной системе (ЦНС), в то время как клетки Шванна миелинизируют часть, которая находится в периферической нервной системе (ПНС). Переход между ЦНС и ПНС происходит на уровне мягкой мозговой оболочки, самого внутреннего и наиболее тонкого слоя менингеальной ткани, окружающего компоненты ЦНС.
Аксон α-МН соединяется с его экстрафузальным мышечным волокном через нервно-мышечное соединение, специальный тип химического синапса, который отличается как по структуре, так и по функциям от химические синапсы, соединяющие нейроны друг с другом. Оба типа синапсов полагаются на нейротрансмиттеры для преобразования электрического сигнала в химический сигнал и обратно. Одно из их различий заключается в том, что синапсы между нейронами обычно используют глутамат или ГАМК в качестве нейромедиаторов, в то время как нервно-мышечные соединения используют исключительно ацетилхолин. Ацетилхолин ощущается никотиновыми рецепторами ацетилхолина на экстрафузальных мышечных волокнах, вызывая их сокращение.
Как и другие двигательные нейроны, α-MN названы в честь свойств их аксонов. Альфа-моторные нейроны имеют аксоны Aα, которые представляют собой большие калибра, сильно миелинизированные волокна, которые быстро проводят потенциалы действия. Напротив, гамма-мотонейроны имеют аксоны Aγ, которые представляют собой тонкие, слегка миелинизированные волокна, которые проводят медленнее.
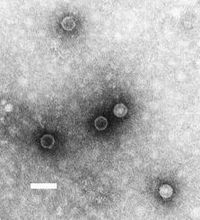 Полиомиелит, вызванный полиовирусом, обнаруженным здесь, связан с избирательной потерей клеток в вентральном роге спинного мозга, где расположены α-МН.
Полиомиелит, вызванный полиовирусом, обнаруженным здесь, связан с избирательной потерей клеток в вентральном роге спинного мозга, где расположены α-МН. Повреждение α-МН является наиболее распространенным типом поражения нижних мотонейронов . Повреждение может быть вызвано, среди прочего, травмой, ишемией и инфекцией. Кроме того, некоторые заболевания связаны с избирательной потерей α-MN. Например, полиомиелит вызывается вирусом, который специфически поражает двигательные нейроны в вентральном роге спинного мозга и убивает их. Боковой амиотропный склероз аналогичным образом связан с избирательной потерей двигательных нейронов.
Паралич - один из наиболее выраженных эффектов повреждения α-МН. Поскольку α-MN обеспечивают единственную иннервацию экстрафузальных мышечных волокон, потеря α-MN эффективно разрывает связь между стволом и спинным мозгом и мышцами, которые они иннервируют. Без этой связи невозможно произвольное и непроизвольное (рефлекторное) управление мышцами. Произвольный мышечный контроль теряется, потому что α-МН передают произвольные сигналы от верхних мотонейронов к мышечным волокнам. Потеря непроизвольного контроля возникает в результате прерывания рефлекторных цепей, таких как тонический рефлекс растяжения. Следствием прерывания рефлекса является снижение мышечного тонуса, в результате чего. Другим следствием является угнетение глубоких сухожильных рефлексов, вызывающих гипорефлексию.
мышечную слабость и атрофию, которые также являются неизбежными последствиями поражений α-МН. Поскольку размер и сила мышц зависят от степени их использования, денервированные мышцы склонны к атрофии. Вторичной причиной мышечной атрофии является то, что денервированные мышцы больше не снабжаются трофическими факторами от α-МН, которые их иннервируют. Поражения альфа-мотонейрона также приводят к аномальным ЭМГ потенциалам (например,) и фасцикуляциям, причем последние представляют собой спонтанные непроизвольные сокращения мышц.
Заболевания, которые нарушают передачу сигналов между α-МН и экстрафузальными мышечными волокнами, а именно заболевания нервно-мышечного соединения, имеют признаки, аналогичные тем, которые возникают при заболевании α-МН. Например, миастения представляет собой аутоиммунное заболевание, которое препятствует передаче сигналов через нервно-мышечное соединение, что приводит к функциональной денервации мышц.