
Войти
| Ретикулярная формация | |
|---|---|
 Венечный разрез моста в его верхней части. (Formatio reticularis обозначен слева.) Венечный разрез моста в его верхней части. (Formatio reticularis обозначен слева.) | |
 Поперечный разрез продолговатого мозга примерно на середине оливы. (Formatio reticularis grisea и formatio reticularis alba отмечены слева.) Поперечный разрез продолговатого мозга примерно на середине оливы. (Formatio reticularis grisea и formatio reticularis alba отмечены слева.) | |
| Подробности | |
| Место нахождения | Мозговой ствол |
| Идентификаторы | |
| латинский | formatio reticularis |
| MeSH | D012154 |
| NeuroNames | 1223 |
| НейроЛекс ID | nlx_143558 |
| TA98 | А14.1.00.021 А14.1.05.403 А14.1.06.327 |
| TA2 | 5367 |
| FMA | 77719 |
| Анатомические термины нейроанатомии [ редактировать в Викиданных ] | |
Ретикулярная формация представляет собой совокупность взаимосвязанных ядер, расположенных по всему мозгу. Он не имеет четкого анатомического определения, поскольку включает нейроны, расположенные в разных частях мозга. Нейроны ретикулярной формации составляют сложный набор сетей в ядре ствола мозга, которые простираются от верхней части среднего мозга до нижней части продолговатого мозга. Ретикулярная формация включает восходящие пути к коре в восходящей ретикулярной активирующей системе ( ARAS) и нисходящие пути к спинному мозгу через ретикулоспинальные пути.
Нейроны ретикулярной формации, особенно нейроны восходящей ретикулярной активирующей системы, играют решающую роль в поддержании поведенческого возбуждения и сознания. Общие функции ретикулярной формации являются модуляторными и премоторными, включая соматический моторный контроль, сердечно-сосудистый контроль, модуляцию боли, сон и сознание, а также привыкание. Модуляторные функции в основном обнаруживаются в ростральном секторе ретикулярной формации, а премоторные функции локализуются в нейронах в более каудальных областях.
Ретикулярная формация разделена на три столбца: ядра шва (медиана), гигантоцеллюлярные ретикулярные ядра (медиальная зона) и парвоцеллюлярные ретикулярные ядра (латеральная зона). Ядра шва являются местом синтеза нейромедиатора серотонина, который играет важную роль в регуляции настроения. Гигантоцеллюлярные ядра участвуют в координации движений. Парвоцеллюлярные ядра регулируют выдох.
Ретикулярная формация необходима для управления некоторыми из основных функций высших организмов и является одной из самых древних филогенетических частей мозга.
 Поперечный разрез нижней части моста, показывающий ретикулярную формацию моста, обозначенный цифрой №9.
Поперечный разрез нижней части моста, показывающий ретикулярную формацию моста, обозначенный цифрой №9. Ретикулярная формация человека состоит из почти 100 ядер мозга и содержит множество выступов в передний мозг, ствол мозга и мозжечок, среди других областей. Он включает ретикулярные ядра, волокна ретикулоталамической проекции, диффузные таламокортикальные проекции, восходящие холинергические проекции, нисходящие нехолинергические проекции и нисходящие ретикулоспинальные проекции. Ретикулярная формация также содержит две основные нейронные подсистемы : восходящую ретикулярную активирующую систему и нисходящие ретикулоспинальные тракты, которые опосредуют различные когнитивные и физиологические процессы. Он был функционально рассечен как сагиттально, так и коронально.
Традиционно ретикулярные ядра делятся на три столбца:
Первоначальная функциональная дифференциация заключалась в разделении на каудальное и ростральное. Это было основано на наблюдении, что повреждение ростральной ретикулярной формации вызывает гиперсомнию в головном мозге кошки. Напротив, поражение более каудальной части ретикулярной формации вызывает бессонницу у кошек. Это исследование привело к мысли, что каудальная часть подавляет ростральную часть ретикулярной формации.
В сагиттальном делении больше морфологических различий. Шва ядра образуют гребень в середине ретикулярной формации, и, непосредственно к его периферии, существует деление называется медиальной ретикулярной формации. Медиальная RF большая, имеет длинные восходящие и нисходящие волокна и окружена латеральной ретикулярной формацией. Латеральная RF находится рядом с двигательными ядрами черепных нервов и в основном опосредует их функцию.
Медиальная ретикулярная формация и латеральная ретикулярная формация - это два столбца ядер с плохо очерченными границами, которые посылают проекции через продолговатый мозг в средний мозг. Ядра можно дифференцировать по функциям, типу клеток и проекциям эфферентных или афферентных нервов. Двигаясь каудально от рострального среднего мозга, на месте рострального моста и среднего мозга, медиальная RF становится менее заметной, а латеральная RF становится более заметной.
По бокам медиальной ретикулярной формации находится ее латеральный кузен, который особенно выражен в ростральном мозговом веществе и каудальном мосту. Из этой области выходят черепные нервы, в том числе очень важный блуждающий нерв. Латеральная RF известна своими ганглиями и областями интернейронов вокруг черепных нервов, которые служат для опосредования их характерных рефлексов и функций.
Ретикулярная формация состоит из более чем 100 небольших нейронных сетей с различными функциями, включая следующие:
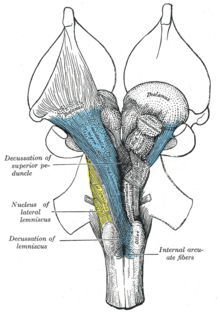 Восходящая ретикулярная активирующая система. Ретикулярная формация отмечена около центра.
Восходящая ретикулярная активирующая система. Ретикулярная формация отмечена около центра. Восходящая ретикулярная активирующая система (ARAS), также известная как модуляторная система экстраталамического контроля или просто ретикулярная активирующая система (RAS), представляет собой набор связанных ядер в мозге позвоночных, которые отвечают за регулирование бодрствования и переходов между сном и бодрствованием. ARAS является частью ретикулярной формации и в основном состоит из различных ядер таламуса и ряда дофаминергических, норадренергических, серотонинергических, гистаминергических, холинергических и глутаматергических ядер мозга.
ARAS состоит из нескольких нервных цепей, соединяющих дорсальную часть заднего среднего мозга и передний мост с корой головного мозга через различные пути, которые проходят через таламус и гипоталамус. ARAS представляет собой совокупность различных ядер - более 20 с каждой стороны в верхнем стволе мозга, мосту, мозговом веществе и заднем гипоталамусе. Нейромедиаторы, которые выделяют эти нейроны, включают дофамин, норэпинефрин, серотонин, гистамин, ацетилхолин и глутамат. Они оказывают корковое влияние через прямые проекции аксонов и косвенные проекции через таламические реле.
Таламический путь состоит в основном из холинергических нейронов в покрышке моста, тогда как гипоталамический путь состоит в основном из нейронов, которые выделяют моноаминовые нейротрансмиттеры, а именно дофамин, норадреналин, серотонин и гистамин. Нейроны, высвобождающие глутамат в ARAS, были идентифицированы гораздо позже по сравнению с моноаминергическими и холинергическими ядрами; глутаматергический компонент ARAS включает одно ядро в гипоталамусе и различные ядра ствола мозга. В Orexin нейроны латерального гипоталамуса иннервируют каждый компонент системы активирующей восходящей ретикулярной и координировать деятельность в рамках всей системы.
| Тип ядра | Соответствующие ядра, которые опосредуют возбуждение | Источники |
|---|---|---|
| Дофаминергические ядра | ||
| Норадренергические ядра |
| |
| Серотонинергические ядра | ||
| Гистаминергические ядра | ||
| Холинергические ядра | ||
| Глутаматергические ядра |
| |
| Ядра таламуса |
ARAS состоит из эволюционно древних областей мозга, которые имеют решающее значение для выживания животного и защищены в неблагоприятные периоды, например, в периоды подавления Totsellreflex, также известного как «животный гипноз». Восходящая ретикулярная активирующая система, которая посылает нейромодуляторные проекции в кору, в основном соединяется с префронтальной корой. Похоже, что связь с моторными областями коры невысока.
Восходящая ретикулярная активирующая система является важным стимулирующим фактором для состояния сознания. Считается, что восходящая система способствует бодрствованию, которое характеризуется корковым и поведенческим возбуждением.
Основная функция ARAS заключается в изменении и усилении таламической и корковой функции, что приводит к десинхронизации электроэнцефалограммы (ЭЭГ). Существуют явные различия в электрической активности мозга в периоды бодрствования и сна: низковольтные быстрые всплески мозговых волн (десинхронизация ЭЭГ) связаны с бодрствованием и быстрым сном (которые электрофизиологически схожи); Медленные волны высокого напряжения обнаруживаются во время медленного сна. Вообще говоря, когда таламические ретрансляционные нейроны находятся в импульсном режиме, ЭЭГ синхронизируется, а когда они находятся в тоническом режиме, она десинхронизируется. Стимуляция ARAS вызывает десинхронизацию ЭЭГ, подавляя медленные корковые волны (0,3–1 Гц), дельта-волны (1–4 Гц) и колебания волны веретена (11–14 Гц), а также стимулируя колебания гамма-диапазона (20–40 Гц)..
Физиологический переход от состояния глубокого сна к бодрствованию обратим и опосредован ARAS. Вентролатеральная преоптическая ядро (VLPO) из гипоталамуса ингибирует нейронные цепи, ответственные за активное состояние, и VLPO способствует активации до начала сна. Во время сна нейроны в ARAS будут иметь гораздо более низкую скорость возбуждения; и наоборот, они будут иметь более высокий уровень активности во время бодрствования. Чтобы мозг мог спать, должно быть снижение восходящей афферентной активности, достигающей коры, путем подавления ARAS.
ARAS также помогает опосредовать переходы от расслабленного бодрствования к периодам повышенного внимания. В ретикулярной формации среднего мозга (MRF) и интраламинарных ядрах таламуса во время выполнения задач, требующих повышенной бдительности и внимания, наблюдается повышенный регионарный кровоток (предположительно, указывающий на повышенную активность нейронов).
Массовые поражения ядер ARAS ствола мозга могут вызывать серьезные изменения уровня сознания (например, кому ). Двустороннее повреждение ретикулярной формации среднего мозга может привести к коме или смерти.
Прямая электрическая стимуляция ARAS вызывает болевые реакции у кошек и вызывает устные сообщения о боли у людей. Восходящая ретикулярная активация у кошек может вызвать мидриаз, который может быть результатом продолжительной боли. Эти результаты предполагают некоторую связь между цепями ARAS и физиологическими путями боли.
Некоторые патологии ARAS могут быть связаны с возрастом, поскольку, по-видимому, наблюдается общее снижение реактивности ARAS с возрастом. Изменения в электрической связи, были предложены для учета некоторых изменений в активности АРАСА: если муфта была вниз регулируется, было бы соответствующее уменьшение синхронизации выше частот (гамма - диапазон). С другой стороны, до регулируемого электрического соединения приведет к увеличению синхронизации быстрых ритмов, которые могли бы привести к увеличению привода сна возбуждения и REM. В частности, нарушение ARAS связано со следующими расстройствами:
Существует несколько потенциальных факторов, которые могут отрицательно повлиять на развитие активирующей системы восходящей ретикулярной системы:
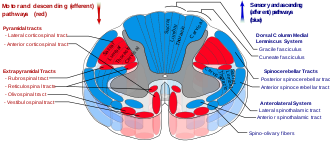 Пути спинного мозга - ретикулоспинальный тракт, обозначенный красным, в центре слева на рисунке.
Пути спинного мозга - ретикулоспинальный тракт, обозначенный красным, в центре слева на рисунке. В ретикулоспинальных трактах, также известные как нисходящие или передних ретикулоспинальные пути, являются экстрапирамидными двигательными трактами, которые происходят из ретикулярной формации в двух трактах, чтобы воздействовать на моторных нейронах, снабжающих туловище и конечностей проксимальных сгибатели и разгибатели. Ретикулоспинальные тракты участвуют в основном в локомоции и контроле позы, хотя у них есть и другие функции. Нисходящие ретикулоспинальные тракты являются одним из четырех основных кортикальных путей к спинному мозгу, отвечающих за опорно-двигательную деятельность. Ретикулоспинальные тракты работают с тремя другими путями, обеспечивая скоординированный контроль движений, включая деликатные манипуляции. Четыре пути можно сгруппировать в два основных системных пути - медиальную систему и латеральную систему. Медиальная система включает ретикулоспинальный путь и вестибулоспинальный путь, и эта система обеспечивает контроль позы. Кортикоспинальные и rubrospinal тракта пути принадлежат к боковой системе, которая обеспечивает контроль тонкого движения.
Этот нисходящий тракт делится на две части: медиальный (или мостовидный) и латеральный (или медуллярный) ретикулоспинальный тракт (MRST и LRST).
Восходящий сенсорный тракт, передающий информацию в противоположном направлении, известен как спиноретикулярный тракт.
Ретикулоспинальные тракты обеспечивают путь, с помощью которого гипоталамус может контролировать симпатический грудопоясничный отток и парасимпатический отток из крестца.
Две основные нисходящие системы, передающие сигналы от ствола головного мозга и мозжечка к спинному мозгу, могут запускать автоматический постуральный ответ для баланса и ориентации: вестибулоспинальные тракты от вестибулярных ядер и ретикулоспинальные тракты от мостов и продолговатого мозга. Поражение этих путей приводит к глубокой атаксии и нестабильности осанки.
Физическое или сосудистое повреждение ствола мозга, разъединяющее красное ядро (средний мозг) и вестибулярные ядра (мосты), может вызвать децеребрационную ригидность, которая имеет неврологический признак повышенного мышечного тонуса и гиперактивных рефлексов растяжения. В ответ на поразительный или болезненный раздражитель обе руки и ноги вытягиваются и поворачиваются внутрь. Причина - тоническая активность латерального вестибулоспинального и ретикулоспинального трактов, стимулирующая двигательные нейроны- разгибатели без угнетения со стороны руброспинального тракта.
Повреждение ствола мозга выше уровня красного ядра может вызвать жесткость декортикации. В ответ на поразительный или болезненный раздражитель руки сгибаются, а ноги вытягиваются. Причина в том, что красное ядро через руброспинальный тракт противодействует возбуждению моторных нейронов-разгибателей из боковых вестибулоспинальных и ретикулоспинальных трактов. Поскольку руброспинальный тракт простирается только до шейного отдела спинного мозга, он в основном воздействует на руки, возбуждая мышцы-сгибатели и подавляя разгибатели, а не ноги.
Повреждение мозгового вещества ниже вестибулярных ядер может вызвать вялый паралич, гипотонию, потерю дыхательного движения и квадриплегию. Отсутствуют рефлексы, напоминающие ранние стадии спинального шока, из-за полной потери активности моторных нейронов, так как больше нет тонической активности, исходящей от латерального вестибулоспинального и ретикулоспинального трактов.
Термин «ретикулярная формация» был придуман в конце 19 века Отто Дейтерсом, что совпадает с учением Рамона-и-Кахала о нейронах. Аллан Хобсон утверждает в своей книге «Пересмотр ретикулярной формации», что это название является этимологическим пережитком упавшей эры теории совокупного поля в нейронных науках. Термин « ретикулум » означает «сетчатая структура», на которую на первый взгляд похож ретикулярная формация. Он был описан как слишком сложный для изучения или как недифференцированная часть мозга, не имеющая никакой организации. Эрик Кандел описывает ретикулярную формацию как организованную аналогично промежуточному серому веществу спинного мозга. Эта хаотичная, рыхлая и запутанная форма организации - вот что оттолкнуло многих исследователей от более глубокого изучения этой конкретной области мозга. Клетки лишены четких ганглиозных границ, но имеют четкую функциональную организацию и различные типы клеток. Термин «ретикулярная формация» больше не используется, за исключением общих слов. Современные ученые обычно называют отдельные ядра, составляющие ретикулярную формацию.
Моруцци и Магун впервые исследовали нейронные компоненты, регулирующие механизмы сна и бодрствования в мозге, в 1949 году. Физиологи предположили, что некая структура глубоко внутри мозга контролирует умственное бодрствование и бдительность. Считалось, что бодрствование зависит только от прямого приема афферентных (сенсорных) стимулов корой головного мозга.
Поскольку прямая электрическая стимуляция мозга может имитировать электрокортикальные реле, Магун использовал этот принцип, чтобы продемонстрировать на двух отдельных участках ствола мозга кошки, как вызвать бодрствование во сне. Сначала он стимулировал восходящие соматические и слуховые пути; во- вторых, из серии «восходящих реле от ретикулярной формации нижнего ствола головного мозга через средний мозг покрышку, subthalamus и гипоталамуса к внутренней капсуле. » Последнее представляло особый интерес, поскольку эта серия реле не соответствовала каким-либо известным анатомическим путям передачи сигнала бодрствования и была придумана как восходящая ретикулярная активирующая система (ARAS).
Затем значение этой недавно идентифицированной релейной системы было оценено путем размещения повреждений в медиальной и латеральной частях передней части среднего мозга. Кошки с мезэнцефалическими перебоями в работе ARAS впадали в глубокий сон и отображали соответствующие мозговые волны. В качестве альтернативы кошки с аналогичным расположением перерывов в восходящих слуховых и соматических путях демонстрировали нормальный сон и бодрствование, и их можно было разбудить с помощью физических раздражителей. Поскольку эти внешние стимулы будут заблокированы на пути к коре из-за прерываний, это указывало на то, что восходящая передача должна проходить через недавно обнаруженный ARAS.
Наконец, Магун зарегистрировал потенциалы в медиальной части ствола мозга и обнаружил, что слуховые стимулы напрямую запускают части ретикулярной активирующей системы. Кроме того, однократная стимуляция седалищного нерва также активировала медиальную ретикулярную формацию, гипоталамус и таламус. Возбуждение ARAS не зависело от дальнейшего распространения сигнала по цепям мозжечка, так как те же результаты были получены после децеребелляции и декортикации. Исследователи предположили, что столб клеток, окружающих ретикулярную формацию среднего мозга, получал данные от всех восходящих трактов ствола головного мозга и передавал эти афференты в кору и, следовательно, регулировал бодрствование.