
Войти
| Мегалозавриды Временной диапазон: средняя – поздняя юра,170–145 млн лет назад PreꞒ Ꞓ О С Д С п Т Дж К пг Н Возможная берриасско - валанжинская запись | |
|---|---|
 | |
| Скелет Torvosaurus tanneri, Музей древней жизни | |
| Научная классификация | |
| Царство: | Анималия |
| Тип: | Хордовые |
| Клада: | Динозаврия |
| Клада: | ящеротазовые |
| Клада: | Тероподы |
| Клада: | † Мегалозаврия |
| Семья: | † Мегалозавриды Хаксли, 1869 г. |
| Типовые виды | |
| † Бакландский мегалозавр Мантелл, 1827 г. | |
| Подгруппы | |
| |
| Синонимы | |
| |
Megalosauridae) — монофилетическое семейство хищных динозавров - теропод, входящее в группу Megalosauroidea. Появившись в средней юре, мегалозавриды были одними из первых крупных радиационных тероподовых динозавров. Они были относительно примитивной группой базальных столбняков, состоящей из двух основных подсемейств, Megalosaurinae и Afrovenatorinae, наряду с базальным родом Eustreptospondylus, нерешенным таксоном, который отличается от обоих подсемейств.
Определяющий мегалозавр, Megalosaurus bucklandii, был впервые назван и описан в 1824 году Уильямом Баклендом после многочисленных находок в Стоунсфилде, Оксфордшир, Великобритания. Мегалозавр был первым официально описанным динозавром и послужил основой для создания клады динозавров. Это также один из крупнейших известных плотоядных динозавров средней юры с наиболее хорошо сохранившейся бедренной костью 805 мм и предполагаемой массой тела около 943 кг. Megalosauridae в основном были признаны европейской группой динозавров на основании окаменелостей, найденных во Франции и Великобритании, но окаменелости показывают, что эта группа также встречается в Северной Америке, Африке, Южной Америке и, возможно, Азии.
Семейство Megalosauridae было впервые определено Томасом Хаксли в 1869 году, однако оно оспаривалось на протяжении всей истории из-за его роли « мусорной корзины » для многих частично описанных динозавров или неопознанных останков. В первые годы палеонтологии большинство крупных теропод были сгруппированы вместе, и до 48 видов были включены в кладу Megalosauria, базальную кладу Megalosauridae. Со временем большинство этих таксонов были помещены в другие клады, и параметры Megalosauridae значительно сузились. Однако остаются некоторые разногласия по поводу того, следует ли считать Megalosauridae отдельной отдельной группой, и динозавры в этом семействе остаются одними из самых проблематичных таксонов во всей Dinosauria. Некоторые палеонтологи, такие как Пол Серено в 2005 году, игнорировали эту группу из-за ее шаткой основы и отсутствия уточненной филогении. Однако недавнее исследование Каррано, Бенсона и Сэмпсона систематически проанализировало всех базальных столбняков и определило, что Megalosauridae должны существовать как отдельное семейство. Как правило, они были тесно связаны с семейством Spinosauridae.
 Масштабное сравнение 3 афровенаторинов
Масштабное сравнение 3 афровенаторинов Как и другие столбняки, мегалозавриды — плотоядные тероподы, характеризующиеся большими размерами и прямохождением на ноги. В частности, мегалозавриды отличаются особенно гигантскими размерами, некоторые представители семейства весят более одной тонны. Со временем есть свидетельства увеличения размеров семьи. Базальные мегалозавриды ранней юры имели меньший размер тела, чем появившиеся в конце средней юры. Из-за этого увеличения размера с течением времени мегалозавриды, по-видимому, следуют схеме увеличения размера, аналогичной другим тероподам гигантского размера, таким как спинозавриды. Эта закономерность следует правилу Коупа, постулату палеонтолога Эдварда Коупа об эволюционном увеличении размеров тела.
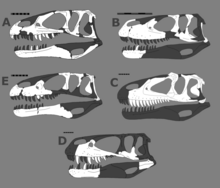 Черепа мегалозавридов. По часовой стрелке сверху слева: Dubreuillosaurus, Torvosaurus, Afrovenator, Megalosaurus, Eustreptospondylus.
Черепа мегалозавридов. По часовой стрелке сверху слева: Dubreuillosaurus, Torvosaurus, Afrovenator, Megalosaurus, Eustreptospondylus. Одна однозначная синапоморфия Megalosauridae - более низкий и длинный череп с соотношением длины к высоте 3: 1. Кроме того, типичная крыша черепа, как правило, гораздо менее орнаментирована, чем у других столбняков, а гребни или рога либо очень маленькие, либо полностью отсутствуют. Мегалозавры также имеют головки бедренных костей с ориентацией на 45 градусов между переднемедиальной и полностью медиальной. Мегалозавриды также определяются следующими уникальными однозначными синапоморфиями:
Megalosaurinae (все мегалозавриды более тесно связаны с Megalosaurus, чем с Afrovenator) характеризуются умеренным (0,5–2,0) соотношением высоты и длины предчелюстной кости ниже уровня ноздрей по сравнению с другими мегалозавридами, которые имеют более низкое соотношение и, следовательно, менее высокую морду. наконечник.
Afrovenatorinae (все мегалозавриды, более близкие к Afrovenator, чем к Megalosaurus) характеризуются квадратным передним краем анторбитальной ямки и широко открытой по средней линии лобково-шадиальной пластинкой.
 Зуб мегалозавра
Зуб мегалозавра Находки зубов часто используются для дифференциации различных теропод и для дальнейшего информирования о кладистической филогении. Морфология зубов и маркеры эволюции зубов склонны к гомоплазии и исчезают или появляются снова на протяжении истории. Однако у мегалозавридов есть несколько специфических состояний протезов, которые отличают их от других базальных теропод. Одно стоматологическое заболевание, присутствующее у Megalosauridae, - это множественные морщины эмали возле килей, острого края или зубчатого ряда зуба. Орнаментированные зубы и хорошо выраженная поверхность эмали также характерны для базальных мегалозавров. Орнамент и хорошо выраженная поверхность появляются у ранних мегалозавридов, но исчезают у производных мегалозавридов, что позволяет предположить, что это состояние со временем было утрачено по мере роста мегалозавридов.
 Бедренная кость мегалозавра
Бедренная кость мегалозавра С самого начала семейства многие экземпляры, найденные в полевых условиях, были ошибочно классифицированы как мегалозавриды. Например, большинство крупных плотоядных, обнаруженных примерно через столетие после того, как Megalosaurus bucklandii назвали Megalosaurus bucklandii, были отнесены к Megalosauridae. Мегалозавр был первой палеонтологической находкой в своем роде, когда Уильям Бакленд обнаружил гигантскую бедренную кость и назвал ее в 1824 году, еще до появления термина «динозаврия». Первоначально вид M. bucklandii был анатомически основан на различных разрозненных костях, найденных в карьерах вокруг деревни Стоунсфилд, Великобритания. Некоторые из этих ранних находок включали правую зубную кость с хорошо сохранившимся зубом, ребра, кости таза и крестцовые позвонки. Когда ранние палеонтологи и исследователи обнаружили больше костей динозавров в окрестностях, они приписали их все M. bucklandii, поскольку на тот момент в истории это был единственный названный и описанный динозавр. Поэтому вид изначально был описан и классифицирован по массе, возможно, несвязанных признаков.
Современная палеонтология впервые начала подходить к проблемному кладистическому разделению мегалозавров в начале 20 века. Фредрих фон Хюне разделил плотоядных теропод, которые все были сгруппированы в широкую категорию мегалозавридов, на два отдельных семейства: более крупных, более гигантских размеров и более мелких, более легко сложенных теропод. Эти две группы были названы Coelurosauria и Pachypodosauria соответственно. Позже Хьюне различал плотоядных и травоядных динозавров в пахиподозаврии, поместив мясоедов в новую группу карнозавров.
По мере того, как было получено больше информации о базальных тероподах и филогенетических характеристиках, современные палеонтологи начали сомневаться в правильном названии этой группы. В 2005 году палеонтолог Пол Серено отказался от использования клады Megalosauridae из-за ее неоднозначной ранней истории в пользу названия Torvosauridae. Сегодня принято считать, что мегалозавриды существовали, по крайней мере, как группа базальных столбняков, из-за того, что у них больше производных таксонов, чем у цератозавров, и что название Megalosauridae должно представлять эту группу. Megalosauridae также имеет приоритет перед Torvosauridae в соответствии с правилами ICZN, регулирующими фамилии.
 Реконструкция Torvosaurus tanneri
Реконструкция Torvosaurus tanneri Мегалозавриды были впервые филогенетически определены в 1869 году Томасом Хаксли, но в течение многих лет использовались как клада «мусорной корзины». В 2002 году Ронан Аллен переопределил кладу после того, как обнаружил на северо-западе Франции полный череп мегалозавра вида Poekilopleuron. Используя признаки, описанные в этом исследовании, Аллен определил Megalosauridae как динозавров, включая Poekilopleuron valesdunesis, теперь известного как Dubreuillosaurus, Torvosaurus, Afrovenator, и всех потомков их общего предка. Аллен также определил два таксона внутри Megalosauridae: Torvosaurinae был определен как все Megalosauridae, более тесно связанные с Torvosaurus, чем с Poekilopleuron и Afrovenator, а Megalosaurinae был определен как все те, которые более тесно связаны с Poekilopleuron. Megalosauridae также подпадает под базальную кладу Megalosauroidea, которая также содержит Spinosauridae. Однако многие таксоны все еще весьма нестабильны и не могут быть помещены в одну кладу с абсолютной уверенностью. Например, Eustreptospondylus и Streptospondylus, хотя они оба определены как Megalosauridae, часто исключаются для создания более стабильных кладограмм, поскольку они не относятся к определенной подгруппе. Представленная здесь кладограмма соответствует Benson (2010) и Benson et al. (2010).
| Мегалозавриды |
| ||||||||||||||||||||||||||||||||||||||||||
 Wiehenvenator, типичный мегалозавр.
Wiehenvenator, типичный мегалозавр. Затем, в 2012 году, Каррано, Бенсон и Сэмпсон провели гораздо более масштабный анализ столбняков и определили мегалозаврию в более широком смысле как кладу, содержащую мегалозавров, спинозавров и всех их потомков. Другими словами, Megalosauria — это группа, состоящая из двух семейств Megalosauridae и его близких родственников Spinosauridae. В рамках этой новой кладограммы Megalosauridae получили новое подсемейство Afrovenatorinae, в которое вошли все мегалозавриды, более тесно связанные с Afrovenator, чем с Megalosaurus.
Каррано, Бенсон и Сэмпсон также включили различных мегалозавридов, которые ранее были исключены из кладограмм в их исследовании 2012 года, таких как Duriavenator и Wiehenvenator в Megalosaurinae и Magnosaurus, Leshansaurus и Piveteausaurus в Afrovenatorinae.
| Мегалозавриды |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Scuirumimus albersodoerferi, небольшой теропод, описанный в 2012 году и сохранивший протоперья, изначально считался молодым мегалозавроидом. Это привело к убеждению, что у мегалозавридов могли быть перья. Однако последующий анализ поместил Sciurumimus в качестве базального целурозавра, и несколько предполагаемых мегалозавроидных синапоморфий, о которых сообщалось в первоначальном описании, являются общими с базальными целурозаврами.
В 2016 году с помощью филогенетического анализа было обнаружено, что Wiehenvenator относится к Megalosauridae как родственный таксон Torvosaurus. Ниже представлена кладограмма, основанная на филогенетическом анализе, проведенном Rauhut et al., Показывающая родство Wiehenvenator.
| Мегалозавриды |
| ||||||||||||||||||||||||||||||||||||||||||||||||
В 2019 году Раухут и Пол описали Asfaltovenator vialidadi, базального аллозавроида, демонстрирующего мозаику примитивных и производных признаков, наблюдаемых у Tetanurae. Их филогенетический анализ показал, что традиционные Megalosauroidea представляют базовый класс карнозавров, парафилетический по отношению к Allosauroida. Это сделало бы Megalosauridae семейством карнозавров.
| Карнозаврия |
| ||||||||||||||||||||||||||||||||||||||||||||||||
 Дорожки мегалозавридов из природного парка Вале-де-Мейос, Серрас-де-Айре и Кандейрос.
Дорожки мегалозавридов из природного парка Вале-де-Мейос, Серрас-де-Айре и Кандейрос. Предполагается, что мегалозавриды являются хищниками или падальщиками, населяющими прибрежную среду. Следы средней юры, которые, как считается, оставили мегалозавриды, были найдены в Вале-де-Мейос в Португалии. В середине юрского периода это место должно было представлять собой приливную равнину, обнажавшуюся во время отлива на краю лагуны. В отличие от большинства прибрежных следов, которые параллельны береговой линии и, вероятно, оставлены мигрирующими животными, следы Вале-де-Мейос были перпендикулярны берегу, причем подавляющее большинство было ориентировано в сторону лагуны. Это указывает на то, что мегалозавриды, оставившие эти следы, подошли к приливной отмели после отступления прилива.
Это указывает на то, что мегалозавриды могли собирать останки морских существ, оставленные отступающими приливами. Другая возможность состоит в том, что мегалозавры были рыбоядными, приближаясь к побережью, чтобы охотиться на рыбу. Спинозавриды, которые были близкими родственниками мегалозавридов, имели многочисленные приспособления к рыбоядной и полуводной жизни, поэтому такой образ жизни подтверждается филогенетическими данными. Зубы акулы, фрагменты хрящей и гастролиты были задокументированы как содержимое желудка Poekilopleuron. И этот род, и Dubreillosaurus были обнаружены в отложениях, также сохранивших корни мангровых зарослей, что является дополнительным свидетельством прибрежной среды обитания. Тем не менее это не исключает возможности того, что мегалозавриды питались и наземной добычей.
 Пангея до разделения на Гондвану и Лавразию
Пангея до разделения на Гондвану и Лавразию Виды, включенные в Megalosauridae, были обнаружены на всех современных континентах, разделенных относительно поровну между местами на суперконтинентах Гондвана и Лавразия. Палеогеографические данные показывают, что мегалозавриды в основном обитали в среднем и позднем юрском периоде, предполагая, что они вымерли на границе юрского и мелового периодов 145 миллионов лет назад. Зубы из берриасско - валанжинской формации Бахада - Колорада в Аргентине позволяют предположить, что эта группа могла сохраняться в раннем меловом периоде в Южной Америке.
Глобальная радиация этих плотоядных теропод происходила в два этапа. Во-первых, радиация произошла во время распада Пангеи в ранней юре, около 200 миллионов лет назад. Когда между суперконтинентом возникло море Тетис, мегалозавроиды распространились на две половины Пангеи. Второй этап радиации произошел в средней и поздней юре, от 174 до 145 миллионов лет назад, у аллозавроидов и целурозавров. Мегалозавриды, по-видимому, вымерли в конце этого периода времени.
Останки мегалозавридов на протяжении всей истории находили в различных частях мира. Например, Megalosauridae содержит самый примитивный зародыш теропод из когда- либо найденных из раннетитонской Португалии 152 миллиона лет назад (млн лет назад). Кроме того, различные открытия окаменелостей мегалозавров были датированы байосом - келловейской Англией и Францией от 168 до 163 млн лет назад, средней юрой в Африке около 170 млн лет назад, позднеюрским Китаем от 163 до 145 млн лет назад и титонским периодом в Северной Америке около 150 млн лет назад. Совсем недавно мегалозавриды были обнаружены в формации Тиурарен в Нигере, что еще раз доказывает, что эти базальные столбняки подверглись глобальному облучению. Зубы из формации Такуарембо позднеюрского периода в Уругвае и формации Тендагуру в Танзании указывают на присутствие крупного мегалозаврина, вероятно, торвозавра.



