
Войти
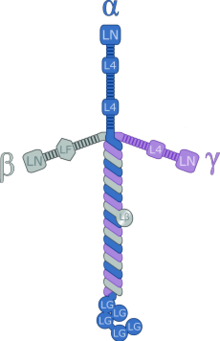 Иллюстрация комплекса ламинина-111, отображающая организацию домена
Иллюстрация комплекса ламинина-111, отображающая организацию домена Ламинины белки с высоким молекулярным весом (от ~ 400 до ~ 900 кДа) внеклеточного матрикса . Они являются основным компонентом базальной пластинки (одного из слоев базальной мембраны ), основы белковой сети для большинства клеток и органов. Ламинины являются важной и биологически активной частью базальной пластинки, влияющей на дифференцировку, миграцию и адгезию клеток.
Ламинины - это гетеротримерные белки, которые содержат α-цепь, β-цепь и γ-цепь., обнаружены в пяти, четырех и трех генетических вариантах соответственно. Молекулы ламинина названы в соответствии с их цепным составом. Таким образом, ламинин-511 содержит цепи α5, β1 и γ1. Четырнадцать других комбинаций цепей были идентифицированы in vivo. Тримерные белки пересекаются, образуя крестообразную структуру, которая может связываться с другими молекулами клеточной мембраны и внеклеточного матрикса. Три более коротких плеча особенно хорошо связываются с другими молекулами ламинина, что позволяет им образовывать листы. Длинное плечо способно связываться с клетками, что помогает прикрепить организованные тканевые клетки к мембране.
Семейство ламининов гликопротеинов является неотъемлемой частью структурной основы почти в каждой ткани организма. Они секретируются и включаются во внеклеточные матрицы, связанные с клетками. Ламинин жизненно важен для поддержания и выживания тканей. Дефектные ламинины могут вызывать неправильное формирование мышц, что приводит к форме мышечной дистрофии, смертельной болезни с образованием пузырей на коже (буллезный эпидермолиз на стыке ) и дефектам почечного фильтра (нефротический синдром ).
Было идентифицировано пятнадцать тримеров ламинина. Ламинины представляют собой комбинации различных альфа-, бета-, и гамма-цепочки.
Ламинины были ранее пронумерованы по мере их открытия, т.е. ламинин-1, ламинин-2, ламинин-3 и т.д., но номенклатура была изменена, чтобы описать, какие цепи присутствуют в каждой изоформе (ламинин-111, ламинин-211 и т.д.). Кроме того, многие ламинины имели общие названия до того, как была введена номенклатура ламининов.
| Старая номенклатура | Старые синонимы | Состав цепи | Новая номенклатура |
|---|---|---|---|
| Ламинин- 1 | EHS ламинин | α1β1γ1 | Ламинин-111 |
| Ламинин-2 | Мерозин | α2β1γ1 | Ламинин- 211 |
| Ламинин-3 | S-ламинин | α1β2γ1 | Ламинин-121 |
| Ламинин-4 | S-мерозин | α2β2γ1 | Ламинин-221 |
| Ламинин-5 / Ламинин-5A | Калинин, эпилигрин, ницитин, ладсин | α3Aβ3γ2 | Ламинин -332 / Ламинин-3A32 |
| Ламинин-5B | α3Bβ3γ2 | Ламинин-3B32 | |
| Ламинин-6 / Ламинин-6A | K-ламинин | α3Aβ1γ1 | Ламинин-311 / Ламинин-3A11 |
| Ламинин-7 / Ламинин-7A | KS-ламинин | α3Aβ2γ1 | Ламинин-321 / Ламинин-3A21 |
| Ламинин-8 | α4β1γ1 | Ламинин-411 | |
| Ламинин-9 | α4β2γ1 | Ламинин-421 | |
| Ламинин-10 | Дрозофила-подобный ламинин | α5β1γ1 | Ламинин-511 |
| Ламинин-11 | α5β2γ1 | Ламинин-521 | |
| Ламинин-12 | α2β1γ3 | Ламинин-213 | |
| Ламинин-14 | α4β2γ3 | Ламинин-423 | |
| α5β2γ2 | Ламинин-522 | ||
| Ламинин-15 | α5β2γ3 | Ламинин-523 |
Ламинины образуют независимые сети и связаны с сетями коллагена типа IV через энтактин, фибронектин и перлекан. Они также связываются с клеточными мембранами через рецепторы интегрина и другие молекулы плазматической мембраны, такие как гликопротеиновый комплекс дистрогликан и гликопротеин лютеранской группы крови.. Посредством этих взаимодействий ламинины вносят критический вклад в прикрепление и дифференцировку клеток, форму и движение клеток, поддержание фенотипа ткани и обеспечение выживания ткани. Некоторые из этих биологических функций ламинина связаны со специфическими аминокислотными последовательностями или фрагментами ламинина. Например, последовательность пептида [GTFALRGDNGDNGQ], которая расположена на альфа-цепи ламинина, способствует адгезии эндотелиальных клеток.
Ламинин альфа4 распространяется в различных ткани, включая периферические нервы, ганглии задних корешков, скелетные мышцы и капилляры; в нервно-мышечном соединении он необходим для синаптической специализации. Было предсказано, что структура домена ламинина-G будет напоминать структуру пентраксина.
Ламинин-111 является основным субстратом, вдоль которого будут проходить аксоны нервов. расти как in vivo, так и in vitro. Например, он прокладывает путь, по которому развивающиеся ганглиозные клетки сетчатки следуют от сетчатки до тектума. Он также часто используется в качестве субстрата в экспериментах с культурами клеток. Присутствие ламинина-1 может влиять на реакцию конуса роста на другие сигналы. Например, ростовые конусы отталкиваются нетрином при выращивании на ламинине-111, но притягиваются к нетрину при выращивании на фибронектине. Этот эффект ламинина-111, вероятно, происходит за счет снижения внутриклеточного циклического АМФ.
Ламинины обогащаются в месте повреждения после повреждения периферического нерва и секретируются Шванновские клетки. Нейроны периферической нервной системы экспрессируют рецепторы интегрина, которые прикрепляются к ламининам и способствуют нейрорегенерации после травмы.
Дисфункциональная структура одного конкретного ламинина, ламинин-211, является причиной одной формы врожденной мышечной дистрофии. Ламинин-211 состоит из цепей α2, β1 и γ1. Распределение ламинина включает мозг и мышечные волокна. В мышцах он связывается с альфа-дистрогликаном и интегрином альфа7 - бета1 через домен G, а другим концом он связывается с внеклеточным матриксом .
Аномальный ламинин-332, который необходим для адгезии эпителиальных клеток к базальной мембране, приводит к состоянию, называемому буллезный эпидермолиз на стыке, которое характеризуется генерализованными волдырями, обильной грануляционной тканью кожи и слизистой оболочки и изъязвлением зубы.
Неисправный ламинин-521 в почечном фильтре вызывает утечку белка в мочу и нефротический синдром.
Некоторые изоформы ламинина участвуют в патофизиологии рака. Большинство транскриптов, несущих внутренний сайт входа в рибосомы (IRES), участвуют в развитии рака через соответствующие белки. Решающее событие в прогрессировании опухоли, называемое переходом эпителия в мезенхиму (EMT), позволяет клеткам карциномы приобретать инвазивные свойства. Недавно сообщалось о трансляционной активации компонента внеклеточного матрикса ламинина B1 (LAMB1) во время EMT, что указывает на механизм, опосредованный IRES. В этом исследовании IRES-активность LamB1 определялась независимыми бицистронными репортерными анализами. Убедительные доказательства исключают влияние криптического промотора или сайтов сплайсинга на управляемую IRES трансляцию LamB1. Кроме того, не было обнаружено никаких других видов мРНК LamB1, возникающих из альтернативных сайтов старта транскрипции или сигналов полиаденилирования, которые объясняют его трансляционный контроль. Картирование 5'-нетранслируемой области LamB1 (UTR) выявило минимальный мотив IRES LamB1 между -293 и -1 перед стартовым кодоном. Примечательно, что аффинная очистка РНК показала, что белок La взаимодействует с LamB1 IRES. Это взаимодействие и его регуляция во время ЭМП были подтверждены иммунопреципитацией рибонуклеопротеидов. Кроме того, La был способен положительно модулировать трансляцию LamB1 IRES. Таким образом, эти данные показывают, что LamB1 IRES активируется путем связывания с La, что приводит к усилению трансляции во время гепатоцеллюлярной EMT.
Вместе с другими основными компонентами ECM, такие как коллагены и фибронектин, ламинины использовались для улучшения культуры клеток млекопитающих, особенно в случае плюрипотентных стволовых клеток, а также некоторых первичных культур клеток, размножение которых на других субстратах может оказаться затруднительным. В продаже имеются два типа ламининов естественного происхождения. Ламинин-111, выделенный из сарком мышей, является одним из популярных типов ламинина, а также смеси ламининов из плаценты человека, которые могут в первую очередь соответствовать ламинину-211, 411 или 511, в зависимости от поставщика. Различные изоформы ламинина практически невозможно выделить из тканей в чистом виде из-за обширного поперечного сшивания и необходимости жестких условий экстракции, таких как протеолитические ферменты или низкий pH, которые вызывают деградацию. Следовательно, рекомбинантные ламинины производятся с 2000 года. Это позволило проверить, могут ли ламинины играть важную роль in vitro, как они играют в организме человека. В 2008 году две группы независимо друг от друга показали, что эмбриональные стволовые клетки мыши можно месяцами выращивать на рекомбинантном ламинине-511. Позже Родин и др. показали, что рекомбинантный ламинин-511 может быть использован для создания полностью свободной от ксено и определенной среды клеточной культуры для культивирования плюрипотентных ES-клеток человека и человеческих iPS-клеток.
| Домен I ламинина | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | Laminin_I | ||||||||
| Pfam | PF06008 | ||||||||
| InterPro | IPR009254 | ||||||||
| |||||||||
| Laminin Domain II | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Symbol | Laminin_II | ||||||||
| Pfam | PF06009 | ||||||||
| InterPro | IPR010307 | ||||||||
| |||||||||
| Ламинин B (домен IV) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | Laminin_B | ||||||||
| Pfam | PF00052 | ||||||||
| InterPro | IPR000034 | ||||||||
| |||||||||
| Ламинин EGF-подобная (домены III и V) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 кристаллическая структура трех последовательных модулей, подобных эпидермальному фактору роста ламинин-типа (le) цепи ламинина гамма1 несущие сайт связывания нидогена кристаллическая структура трех последовательных модулей, подобных эпидермальному фактору роста ламинин-типа (le) цепи ламинина гамма1 несущие сайт связывания нидогена | |||||||||
| Идентификаторы | |||||||||
| Символ | Laminin_EGF | ||||||||
| Pfam | PF00053 | ||||||||
| Pfam клан | CL0001 | ||||||||
| InterPro | IPR002049 | ||||||||
| PROSITE | PDOC00021 | ||||||||
| SCOPe | 1tle / SUPFAM | ||||||||
| |||||||||
| домен ламинина G | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 пара доменов lg4-5 ламинина альфа 2 цепи, мутант по сайту ca1 пара доменов lg4-5 ламинина альфа 2 цепи, мутант по сайту ca1 | |||||||||
| идентификаторы | |||||||||
| символ | ламинин_G_1 | ||||||||
| Pfam | PF00054 | ||||||||
| Pfam clan | CL0004 | ||||||||
| InterPro | IPR012679 | ||||||||
| SCOPe | 1qu0 / SUPFAM | ||||||||
| |||||||||
| домен ламинина G | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 структура лиганд-связывающего домена нейрексина 1beta: регуляция функции lns-домена посредством альтернативного сплайсинга структура лиганд-связывающего домена нейрексина 1beta: регуляция функции lns-домена посредством альтернативного сплайсинга | |||||||||
| Идентификаторы | |||||||||
| символ | Laminin_G_2 | ||||||||
| Pfam | PF02210 | ||||||||
| Pfam клан | CL0004 | ||||||||
| InterPro | IPR012680 | ||||||||
| SMART | TSPN | ||||||||
| |||||||||
| N-концевой ламинин (домен VI) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | Laminin_N | ||||||||
| Pfam | PF00055 | ||||||||
| Pfam clan | CL0202 | ||||||||
| InterPro | IPR008211 | ||||||||
| SMART | LamNT | ||||||||
| SCOPe | 1klo / SUPFAM | ||||||||
| |||||||||
Ламинины содержат несколько консервативных белковых доменов.
Ламинины представляют собой тримерные молекулы; ламинин-1 является тримером альфа1 бета1 гамма1 . Было высказано предположение, что домены I и II ламинина A, B1 и B2 могут объединяться, образуя тройную спиральную спиральную структуру.
Домен ламинина B (также известный как домен IV) представляет собой внеклеточный модуль с неизвестной функцией. Он обнаружен в ряде различных белков, включая гепарансульфат протеогликан из базальной мембраны, ламинин-подобный белок из Caenorhabditis elegans и ламинин. Домен ламинина IV не обнаруживается в коротких цепях ламинина (альфа4 или бета3).
Помимо различных типов глобулярных доменов каждая субъединица ламинина содержит в своей первой половине последовательные повторы примерно 60 аминокислот длиной, которые включают восемь консервативных цистеинов. Третичная структура этого домена отдаленно похожа по своему N-концу на структуру EGF-подобного модуля. Он также известен как «LE» или «EGF-подобный домен ламининового типа». Число копий EGF-подобного домена ламинина в различных формах ламининов сильно варьирует; найдено от 3 до 22 экземпляров. Было показано, что в цепи гамма-1 ламинина мыши седьмой LE-домен является единственным, который связывает с высоким сродством с нидогеном. Сайты связывания расположены на поверхности внутри петель C1-C3 и C5-C6. Длинные последовательные множества ламининовых EGF-подобных доменов в ламининах образуют стержневидные элементы с ограниченной гибкостью, которые определяют интервалы в образовании ламининовых сетей базальных мембран.
Глобулярный ламинин (G) домен, также известный как домен LNS (ламинин-альфа, нейрексин и глобулин, связывающий половые гормоны), имеет в среднем 177 аминокислот в длину и может быть найден от одной до шести копий в различных члены семейства ламининов, а также в большом количестве других внеклеточных белков. Например, все альфа-цепи ламинина имеют пять доменов ламинина G, все белки семейства коллаген имеют один домен ламинина G, белки CNTNAP имеют четыре домена ламинина G, а нейрексин 1 и 2 каждый содержат шесть доменов ламинина G.. В среднем примерно четверть белков, которые содержат домены ламинина G, захватываются этими доменами ламинина G сами по себе. Наименьший домен ламинина G можно найти в одном из белков коллагена (COL24A1; 77 AA), а самый большой домен - в TSPEAR (219 AA).
Точная функция доменов ламинина G осталась неуловимой, и различные связывающие функции были приписаны различным модулям ламинина G. Например, каждая из цепей ламинина альфа1 и альфа2 имеет пять С-концевых доменов ламинина G, из которых только домены LG4 и LG5 содержат сайтов связывания для гепарина, сульфатидов и рецептор клеточной поверхности дистрогликан. белки, содержащие ламинин G, по-видимому, играют самые разные роли в адгезии клеток, передаче сигналов, миграции, сборке и дифференциация.
Сборка базальной мембраны представляет собой кооперативный процесс, в котором ламинины полимеризуются через свой N-концевой домен (LN или домен VI) и прикрепляются к ячейка через их G-домены. Нетрины могут также связываться с этой сетью через гетеротипные взаимодействия домена LN. Это приводит к передаче сигналов клеток через интегрины и дистрогликан (и, возможно, другие рецепторы), рекрутируемых на прикрепленный ламинин. Эта зависимая от LN-домена самосборка считается решающей для целостности базальных мембран, что подчеркивается формами мышечной дистрофии, содержащих делецию LN-модуля из альфа 2 цепи ламинина. N-концевой домен ламинина обнаружен во всех субъединицах ламинина и нетрина, кроме ламинина альфа 3A, альфа 4 и гамма 2.
LAMA1 ; LAMA2 ; LAMA3 ; LAMA4 ; LAMA5 ;
LAMA1 ; LAMA2 ; LAMA3 ; LAMA4 ; LAMA5 ;
HSPG2 ; LAMA1 ; LAMA2 ; LAMA3 ; LAMA5 ; LAMC1 ; LAMC2 ; LAMC3 ;
AGRIN ; ATRN ; ; CELSR1 ; CELSR2 ; CELSR3 ; CRELD1 ; HSPG2 ; LAMA1 ; LAMA2 ; LAMA3 ; LAMA4 ; LAMA5 ; LAMB1 ; LAMB2 ; LAMB3 ; LAMB4 ; LAMC1 ; LAMC2 ; LAMC3 ; MEGF10 ; ; MEGF6 ; MEGF8 ; ; ; NTN1 ; ; ; NTNG1 ; NTNG2 ; RESDA1 ; ; ; ; STAB1 ; USH2A ;
AGRIN ; CELSR1 ; CELSR2 ; CELSR3 ; CNTNAP1 ; CNTNAP2 ; ; ; CNTNAP4 ; ; COL11A1 ; COL11A2 ; COL12A1 ; COL14A1 ; COL15A1 ; COL16A1 ; COL18A1 ; COL19A1 ; COL20A1 ; COL21A1 ; COL22A1 ; COL24A1 ; COL27A1 ; COL5A1 ; COL5A3 ; COL9A1 ; CRB1 ; CRB2 ; CSPG4 ; ; ; FAT ; ; ; FAT4 ; ГАЗ6 ; HSPG2 ; LAMA1 ; LAMA2 ; LAMA3 ; LAMA4 ; LAMA5 ; NELL1 ; NELL2 ; NRXN1 ; NRXN2 ; NRXN3 ; PROS1 ; SLIT1 ; SLIT2 ; SLIT3 ; КОПЬЕ ; THBS1 ; THBS2 ; THBS3 ; THBS4 ; USH2A ;
LAMA1 ; LAMA2 ; LAMA3 ; LAMA5 ; LAMB1 ; LAMB2 ; LAMB3 ; LAMB4 ; LAMC1 ; LAMC3 ; NTN1 ; ; ; NTNG1 ; NTNG2 ; USH2A ;
.
Эта статья включает текст из общественного достояния Pfam и InterPro :