Ферменты, удаляющие метиловые (CH3-) группы из нуклеиновых кислот
Деметилазы являются ферментами, которые удаляют группы метил (CH 3 -) с нуклеиновых кислот, белков (в частности, гистонов ) и других молекул. Ферменты деметилазы важны в механизмах эпигенетической модификации. Белки деметилазы изменяют регуляцию транскрипции генома, контролируя уровни метилирования, которые возникают в ДНК и гистонах, и, в свою очередь, регулируют хроматин состояния в конкретных локусах гена внутри организмов.
Содержание
- 1 Окислительное деметилирование
- 1.1 Деметилирование гистонов
- 2 Деметилирование сложного эфира
- 3 См. Также
- 4 Ссылки
Окислительное деметилирование
Деметилирование гистонов

Механизмы деметилирования лизина лизиндеметилазой 1A (KDM1A) и гистоновыми деметилазами, содержащими JmjC-домен (JHDM). Оба механизма включают окисление метильной группы (с помощью
FAD или
α-кетоглутарата и молекулярного кислорода в качестве окислителей) с последующим удалением
формальдегида. KDM1A требует наличия атома водорода (зеленого цвета), связанного с метилированным азотом для проявления своей деметилазной активности, и поэтому может деметилировать только моно- и диметилированные лизины.
В течение многих лет метилирование гистонов считалось необратимым из-за к тому, что период полураспада метилирования гистонов был примерно равен периоду полужизни самих гистонов. В 2004 году Shi et al. опубликовали свое открытие гистон-деметилазы LSD1 (позже классифицированной как KDM1A ), гомолога ядерной аминоксидазы. С тех пор было обнаружено еще много гистоновых деметилаз. В зависимости от их механизмов существует два основных класса гистоновых деметилаз: флавинадениндинуклеотид (FAD) -зависимый амин оксидаза и Fe (II) и α-кетоглутарат-зависимая гидроксилаза. Оба действуют путем гидроксилирования метильной группы с последующей диссоциацией формальдегида. Деметилирование имеет значение для эпигенетики..
Белки гистон-деметилазы имеют множество доменов, которые выполняют разные функции. Эти функции включают связывание с гистоном (или иногда ДНК на нуклеосоме ), распознавание правильного метилированного субстрата аминокислоты и катализирование реакции, а также связывание кофакторов. Кофакторы включают: альфа-кетоглутарат (JmjC-домен, содержащий деметилазы), CoREST (LSD), FAD, Fe (II) или NOG (N-оксалилглицин). Домены включают:
- SWIRM1 (домен Swi3, Rsc и Moira): предполагаемый якорный сайт для молекул гистонов; обнаружен в нескольких комплексах, модифицирующих хроматин ; способствует белку деметилазы и кофактору CoREST (связывание с нуклеосомным субстратом)
- Jumonji (N / C-концевые домены): домен связывания ключевых кофакторов, таких как альфа-кетоглутарат ; соединены бета-шпилькой / смешанным доменом
- PHD-finger: гидрофобная клетка остатков, которая связывает метилированные пептиды ; играет ключевую роль в распознавании и селективности в отношении остатков метилированных гистонов
- Цинк-палец : ДНК-связывающий домен
- Аминоксидазный домен : каталитический активный сайт белков LSD; больше, чем родственные белки, чтобы соответствовать размеру гистонового хвоста

Структура JmJDA (координаты из файла PDB: 2UXX); Некоторые домены сверху выделены: JmJ (N-конец, красный; C-конец, желтый), домен цинкового пальца (светло-фиолетовый), бета-шпилька (светло-голубой) и линкер смешанного домена (зеленый).

Структура KDM1A (координаты из файла PDB: 2Z5U)
Существует несколько семейств гистоновых деметилаз, которые действуют на разные субстраты и играют разные роли в клеточной функции. Был разработан код для обозначения субстрата гистоновой деметилазы. Субстрат сначала определяется субъединицей гистона (H1, H2A, H2B, H3, H4), а затем однобуквенным обозначением и номером аминокислоты, которая метилирована. Наконец, уровень метилирования иногда отмечается добавлением «me #» с номерами 1, 2 и 3 для монометилированных, диметилированных и триметилированных субстратов соответственно. Например, H3K9me2 - это гистон H3 с диметилированным лизином в девятом положении.
- KDM1
- Семейство KDM1 включает KDM1A и KDM1B. KDM1A (также называемый LSD1 / AOF2 / BHC110) может действовать на моно- и диметилированный H3K4 и H3K9, а KDM1B (также называемый LSD2 / AOF1) действует только на моно- и диметилированный H3K4. Эти ферменты могут играть решающую роль в эмбриогенезе и тканеспецифической дифференцировке, а также в росте ооцитов. KDM1A была первой открытой деметилазой, и поэтому она была изучена наиболее широко.
- Делеция гена KDM1A может оказывать влияние на рост и дифференцировку эмбриональных стволовых клеток и может привести к эмбриональному летальность у мышей с нокаутом, которые не продуцируют продукт гена KDM1A. Считается, что KDM1A также играет роль в развитии рака, поскольку более неблагоприятные исходы могут быть коррелированы с более высокой экспрессией этого гена. Следовательно, ингибирование KDM1A может быть возможным лечением рака. KDM1A имеет множество различных партнеров по связыванию, которые могут быть необходимы для его активности деметилирования.
- KDM1B, однако, в основном участвует в развитии ооцитов. Делеция этого гена приводит к материнскому эффекту летальности у мышей. Ортологи KDM1 в D. melanogaster и C. elegans, похоже, функционирует аналогично KDM1B, а не KDM1A.
- KDM2
- Семейство KDM2 включает KDM2A и KDM2B. KDM2A (также известный как JHDM1A / FBXL11) может действовать на моно- и диметилированный H3K36 и триметилированный H3K4. KDM2B (также называемый JHDM1B / FBXL10) действует только на моно- и диметилированный H3K36. KDM2A играет роль либо в стимулировании, либо в ингибировании функции опухоли, а KDM2B играет роль в онкогенезе.
- У многих эукариот белок KDM2A содержит домен цинкового пальца CXXC, способный связывать неметилированные CpG-островки. В настоящее время считается, что белки KDM2A могут связываться со многими регуляторными элементами генов без помощи специфичных для последовательности факторов транскрипции. Хотя роль KDM2 в дифференцировке развития эукариот по-прежнему остается загадкой, было показано, что как KDM2A, так и KDM2B играют роль в росте и подавлении опухолей. Было показано, что KDM2B сверхэкспрессируется в человеческих лимфомах и аденокарциномах ; однако при раке простаты и глиобластомах наблюдается пониженная экспрессия как KDM2A, так и KDM2B. Кроме того, было показано, что KDM2B предотвращает старение в некоторых клетках посредством эктопической экспрессии, что дополнительно указывает на его потенциал как онкогена.
- KDM3
- Семейство KDM3 включает KDM3A, KDM3B и JMJD1C. KDM3A (также называемый JHDM2A / JMJD1A / TSGA) может действовать на моно- и диметилированный H3K9. Подложки для KDM3B (также обозначаемые как JHDM2B / JMJD1B) и JMJD1C (также обозначаемые как JHDM2C / TRIP8) неизвестны. KDM3A играет роль в сперматогенезе и метаболических функциях; роль KDM3B и JMJD1C неизвестна.
- Исследования нокдауна KDM3A на мышах, где мышь продуцирует пониженные уровни KDM3A, привели к мужскому бесплодию и появлению ожирения у взрослых. Дополнительные исследования показали, что KDM3A может играть роль в регуляции генов, зависимых от андрогенных рецепторов, а также генов, участвующих в плюрипотентности, что указывает на потенциальную роль KDM3A в онкогенезе.
- KDM4
- KDM4 Семейство включает KDM4A, KDM4B, KDM4C и KDM4D. Они также называются JMDM3A / JMJD2A, JMDM3B / JMJD2B, JMDM3C / JMJD2C и JMDM3D / JMJD2D соответственно. Эти ферменты могут действовать на ди- и триметилированные H3K9, H3K36, H1K26. KDM4B и KDM4C играют роль в туморогенезе, и роль KDM4D неизвестна.
- Семейство белков KDM4 связано со злокачественной трансформацией. В частности, амплификация KDM4C была документально подтверждена при плоскоклеточном раке пищевода, медуллобластоме и раке груди; Амплификация KDM4B также была обнаружена в медуллобластомах. Другие данные по экспрессии генов также предполагают избыточную экспрессию KDM4A, KDM4B и KDM4C при раке простаты.
- KDM5
- Семейство KDM5 включает KDM5A, KDM5B, KDM5C и KDM5D. Они также обозначаются как JARID1A / RBP2, JARID1B / PLU-1, JARID1C / SMCX и JARID1D / SMCY соответственно. Эти ферменты могут действовать на ди- и триметилированный H3K4.
- Семейство белков KDM5, по-видимому, играет ключевые функции развития. Делеция домена JmjC ретинобластомного связывающего белка, родственного 2 (RBR-2) в C. elegans выражают дефекты формирования вульвы. Мутации в домене JmjC у Drosophila вызывают либо летальные эффекты для личинок, либо многие дефекты развития у выживших.
- KDM5A в системах клеточных культур также продемонстрировал связь с регуляцией дифференцировки, митохондрий функция, клеточный цикл прогрессирование. KDM5B и KDM5C также показали взаимодействие с белками PcG, которые участвуют в репрессии транскрипции. Мутации KDM5C (обнаруженные на X-хромосоме) также были обнаружены у пациентов с X-связанной умственной отсталостью. Истощение гомологов KDM5C в D. rerio выявили дефекты формирования паттерна мозга и гибель нейронных клеток.
- KDM6
- Семейство KDM6 включает KDM6A, KDM6B и UTY. KDM6A (также обозначаемый как UTX) и KDM6B (также обозначаемый как JMJD3) действуют на ди- и триметилированный H3K27 и играют роль в развитии; субстрат и роль UTY неизвестны. В целом, и KDM6A, и KDM6B обладают опухолевыми характеристиками. Нокдауны KDM6A в фибробластах приводят к немедленному увеличению популяции фибробластов. KDM6B, экспрессируемый в фибробластах, индуцирует онкогены пути RAS_RAF. Делеции и точечные мутации KDM6A были идентифицированы как одна из причин синдрома Кабуки, врожденного нарушения, приводящего к умственной отсталости.
- Были предложены другие возможные роли KDM6B. В частности, в одном исследовании мутирующие гомологи KDM6B нарушали развитие гонад у C.elegans. Другие исследования показали, что экспрессия KDM6B активируется в активированных макрофагах и динамически экспрессируется во время дифференцировки стволовых клеток.
- . С другой стороны, истощение гомологов KDM6A в D. rerio показали сниженную экспрессию генов HOX, которые играют роль в регуляции формирования паттерна тела во время развития. В исследованиях на млекопитающих было показано, что KDM6A также регулирует гены HOX.
Деметилирование сложного эфира

Карикатурное изображение молекулярной структуры белка, зарегистрированное с помощью кода 1A2O pdb.
Другим примером деметилазы является белок -глутаматметилэстераза, также известная как белок CheB (EC 3.1.1.61), который деметилирует MCP (m этил-акцептирующий c гемотаксис p ротеины) за счет гидролиза сложноэфирных связей карбоновых кислот. Ассоциация рецептора хемотаксиса с агонистом приводит к фосфорилированию CheB. Фосфорилирование белка CheB усиливает его каталитическую активность деметилирования МСР, что приводит к адаптации клетки к раздражителям окружающей среды. MCP реагируют на внеклеточные аттрактанты и репелленты у таких бактерий, как E. coli в регуляции хемотаксиса. CheB более конкретно называют метилэстеразой, так как он удаляет метильные группы из остатков метилглутамата, расположенных на MCP, посредством гидролиза, с образованием глутамата, сопровождаемого высвобождением метанол.
CheB представляет особый интерес для исследователей, поскольку он может быть терапевтической мишенью для уменьшения распространения бактериальных инфекций.
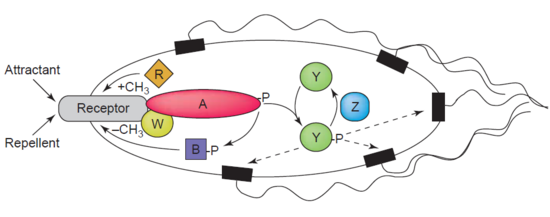
Передача сигналов хемотаксиса. Хемоаттрактанты или репелленты воспринимаются трансмембранными рецепторами. Обратите внимание на роль CheB (B) в деметилировании рецепторов MCP.
См. Также
Ссылки

 Механизмы деметилирования лизина лизиндеметилазой 1A (KDM1A) и гистоновыми деметилазами, содержащими JmjC-домен (JHDM). Оба механизма включают окисление метильной группы (с помощью FAD или α-кетоглутарата и молекулярного кислорода в качестве окислителей) с последующим удалением формальдегида. KDM1A требует наличия атома водорода (зеленого цвета), связанного с метилированным азотом для проявления своей деметилазной активности, и поэтому может деметилировать только моно- и диметилированные лизины.
Механизмы деметилирования лизина лизиндеметилазой 1A (KDM1A) и гистоновыми деметилазами, содержащими JmjC-домен (JHDM). Оба механизма включают окисление метильной группы (с помощью FAD или α-кетоглутарата и молекулярного кислорода в качестве окислителей) с последующим удалением формальдегида. KDM1A требует наличия атома водорода (зеленого цвета), связанного с метилированным азотом для проявления своей деметилазной активности, и поэтому может деметилировать только моно- и диметилированные лизины.  Структура JmJDA (координаты из файла PDB: 2UXX); Некоторые домены сверху выделены: JmJ (N-конец, красный; C-конец, желтый), домен цинкового пальца (светло-фиолетовый), бета-шпилька (светло-голубой) и линкер смешанного домена (зеленый).
Структура JmJDA (координаты из файла PDB: 2UXX); Некоторые домены сверху выделены: JmJ (N-конец, красный; C-конец, желтый), домен цинкового пальца (светло-фиолетовый), бета-шпилька (светло-голубой) и линкер смешанного домена (зеленый).  Структура KDM1A (координаты из файла PDB: 2Z5U)
Структура KDM1A (координаты из файла PDB: 2Z5U)  Карикатурное изображение молекулярной структуры белка, зарегистрированное с помощью кода 1A2O pdb.
Карикатурное изображение молекулярной структуры белка, зарегистрированное с помощью кода 1A2O pdb.  Передача сигналов хемотаксиса. Хемоаттрактанты или репелленты воспринимаются трансмембранными рецепторами. Обратите внимание на роль CheB (B) в деметилировании рецепторов MCP.
Передача сигналов хемотаксиса. Хемоаттрактанты или репелленты воспринимаются трансмембранными рецепторами. Обратите внимание на роль CheB (B) в деметилировании рецепторов MCP.