
Войти
Аксональный транспорт, также называемый аксоплазматический транспорт или аксоплазматический поток, клеточный процесс, ответственный за движение митохондрий, липидов, синаптических пузырьков, белков и других органеллы в и от нейрона клеточного тела через цитоплазму его аксона, называемого аксоплазма. Поскольку некоторые аксоны имеют длину порядка метров, нейроны не могут полагаться на диффузию, чтобы переносить продукты ядра и органелл к концам своих аксонов. Аксональный транспорт также отвечает за перемещение молекул, предназначенных для деградации, от аксона обратно в тело клетки, где они расщепляются лизосомами.
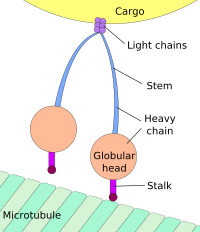 Динеин, моторным белком, ответственным за ретроградный аксональный транспорт, переносит везикулы и другие клеточные продукты по направлению к клеточным телам нейронов. Его легкие цепи связывают груз, а его глобулярные участки головки связывают микротрубочку, «медленно двигаясь» по ней.
Динеин, моторным белком, ответственным за ретроградный аксональный транспорт, переносит везикулы и другие клеточные продукты по направлению к клеточным телам нейронов. Его легкие цепи связывают груз, а его глобулярные участки головки связывают микротрубочку, «медленно двигаясь» по ней. Движение к телу клетки называется ретроградным транспортом, а движение к синапсу называется антероградным переносом.
 Кинезин, идущий по микротрубочке. Это молекулярная биологическая машина, которая использует динамику белковых доменов на наномасштабах
Кинезин, идущий по микротрубочке. Это молекулярная биологическая машина, которая использует динамику белковых доменов на наномасштабах . Подавляющее большинство аксональных белков синтезируется в теле нейрональной клетки и транспортируется по аксонам. Некоторая трансляция мРНК была продемонстрирована в аксонах. Транспорт аксонов происходит на протяжении всей жизни нейрона и необходим для его роста и выживания. Микротрубочки (сделанные из тубулина ) проходят вдоль аксона и обеспечивают основные цитоскелетные «дорожки» для транспортировки. Кинезин и динеин - это моторные белки, которые перемещают грузы в антероградном (вперед от сомы к кончику аксона) и ретроградном (назад) к соме (телу клетки)) соответственно. Моторные белки связываются и транспортируют несколько различных грузов, включая митохондрии, цитоскелет полимеры, аутофагосомы и синаптические везикулы, содержащие нейротрансмиттеры.
Аксональный транспорт может быть быстрым или медленным, а также антероградным (от тела клетки) или ретроградным (транспортирует материалы от аксона к телу клетки).
Везикулярные грузы перемещаются относительно быстро (50–400 мм / день), тогда как транспортировка растворимых (цитозольных) и цитоскелетных белков занимает гораздо больше времени (перемещение менее 8 мм / день). день). Базовый механизм быстрого аксонального транспорта был понят на протяжении десятилетий, но механизм медленного аксонального транспорта стал ясен только недавно, в результате передовых методов визуализации. Методы флуоресцентного мечения (например, флуоресцентная микроскопия ) сделали возможным прямую визуализацию транспорта в живых нейронах. (См. Также: Антероградное отслеживание.)
Недавние исследования показали, что движение цитоскелетных «медленных» грузов на самом деле происходит быстро, но в отличие от быстрых грузов, они часто останавливаются, что делает общую скорость транзита намного медленнее. Этот механизм известен как модель медленного аксонального транспорта «Stop and Go» и был широко подтвержден для транспорта нейрофиламента белка цитоскелета. Движение растворимых (цитозольных) грузов является более сложным, но, по-видимому, имеет аналогичную основу, когда растворимые белки организуются в мультибелковые комплексы, которые затем передаются посредством временных взаимодействий с более быстро движущимися грузами, перемещающимися в быстром аксональном транспорте. Аналогия - разница в тарифах на проезд между местными и скоростными поездами метро. Хотя оба типа поездов движутся между станциями с одинаковой скоростью, местному поезду требуется гораздо больше времени, чтобы добраться до конца линии, потому что он останавливается на каждой станции, тогда как экспресс делает всего несколько остановок по пути.
Антероградный (также называемый «ортоградным») перенос - это движение молекул / органелл наружу от тела клетки (также называемого сома ) к синапсу или клеточной мембране.
Антероградное движение отдельных грузов (в транспортных пузырьках ) как быстрых, так и медленных компонентов вдоль микротрубочек опосредуется кинезинами. Некоторые кинезины участвуют в медленном транспорте, хотя механизм генерации "пауз" при прохождении медленных компонентов груза все еще неизвестен.
Существует два класса медленного антероградного транспорта: медленный компонент a (SCa), который переносит в основном микротрубочки и нейрофиламенты со скоростью 0,1-1 миллиметра в день, и медленный компонент b (SCb), несущий более 200 различных белков и актина со скоростью до 6 миллиметров в день. Медленный компонент b, который также несет актин, транспортируется в аксонах клеток сетчатки со скоростью 2-3 миллиметра в день.
Во время латентной реактивации вирус простого герпеса (HSV) входит в свой литический цикл и использует механизмы антероградного транспорта для миграции из ганглиев дорсальных корешков нейронов кожи или слизистой оболочки, на которые он впоследствии влияет.
Рецептор карго для двигателей антероградного транспорта, кинезины, был идентифицирован как белок-предшественник амилоида (АРР), родительский белок, который производит старческий бляшки, обнаруженные при болезни Альцгеймера. Пептид из 15 аминокислот на карбоксильном конце цитоплазмы APP связывается с высоким сродством с обычным кинезином-1 и опосредует транспорт экзогенного груза в гигантском аксоне кальмара.
Марганец, контрастное вещество для T 1 МРТ, перемещается антероградным транспортом после стереотаксической инъекции в мозг экспериментальных животных и, таким образом, выявляет схему с помощью МРТ всего мозга у живых животных, как это было впервые сделано Робией Паутлер, Элейн Беэр и Расс Джейкобс. Исследования на мышах, лишенных легкой цепи кинезина-1, показали, что Mn перемещается посредством транспорта на основе кинезина в зрительном нерве и в головном мозге. Транспорт как в проекциях гиппокампа, так и в зрительном нерве также зависит от APP. Транспорт из гиппокампа в передний мозг снижается при старении, а место назначения изменяется из-за наличия бляшек при болезни Альцгеймера.
Ретроградный транспорт перемещает молекулы / органеллы от концов аксона к тело клетки. Ретроградный аксональный транспорт опосредуется цитоплазматическим динеином и используется, например, для отправки химических сообщений и продуктов эндоцитоза, направляемых в эндолизосомы от аксона обратно в клетку. Работая со средней скоростью in vivo примерно 2 мкм / сек, быстрый ретроградный транспорт может покрывать 10-20 см в день.
Быстрый ретроградный транспорт возвращает использованные синаптические везикулы и другие материалы в сому и информирует сому об условиях на окончаниях аксонов. Ретроградный транспорт переносит сигналы выживания из синапса обратно в тело клетки, такие как TRK, рецептор фактора роста нервов. Некоторые патогены используют этот процесс для вторжения в нервную систему. Они входят в дистальные кончики аксона и перемещаются к соме ретроградным транспортом. Примеры включают столбнячный токсин и вирусы простого герпеса, бешенства и полиомиелита. При таких инфекциях задержка между инфицированием и появлением симптомов соответствует времени, необходимому для того, чтобы патогены достигли соматических клеток. Вирус простого герпеса перемещается по аксонам в обоих направлениях в зависимости от его жизненного цикла, при этом ретроградный транспорт доминирует полярность для поступающих капсидов.
Когда аксональный транспорт ингибируется или прерывается, нормальная физиология становится патофизиологией, и может возникнуть скопление аксоплазмы, называемое аксональным сфероидом . Поскольку транспорт аксонов может быть нарушен множеством способов, аксональные сфероиды можно увидеть при многих различных классах заболеваний, включая генетические, травматические, ишемические, инфекционные, токсические, дегенеративные и так называемые лейкоэнцефалопатии. Несколько редких нейродегенеративных заболеваний связаны с генетическими мутациями в двигательных белках, кинезине и динеине, и в этих случаях вполне вероятно, что аксональный транспорт является ключевым игроком в опосредовании патологии. Дисфункциональный аксональный транспорт также связан со спорадическими (распространенными) формами нейродегенеративных заболеваний, таких как болезнь Альцгеймера и болезнь Паркинсона. Это в основном связано с многочисленными наблюдениями, что большие скопления аксонов неизменно наблюдаются в пораженных нейронах, и что гены, о которых известно, что они играют роль в семейных формах этих заболеваний, также предположительно играют роль в нормальном аксональном транспорте. Однако прямых доказательств участия аксонального транспорта в последних заболеваниях мало, и другие механизмы (такие как прямая синаптотоксичность) могут быть более актуальными.
Остановка потока аксоплазмы на краю ишемической области при сосудистых ретинопатиях приводит к набуханию нервных волокон, которое приводит к образованию мягких экссудатов или ватных пятен.
Поскольку аксон зависит от аксоплазматического транспорта жизненно важных белков и материалов, повреждение, такое как диффузное повреждение аксона, которое прерывает транспортировку, вызовет дегенерацию дистального аксона в процесс называется валлеровским вырождением. Противораковые препараты, которые препятствуют росту злокачественных опухолей, изменяя микротрубочки (которые необходимы для деления клеток ), повреждают нервы, поскольку микротрубочки необходимы для транспорта аксонов.
вирус бешенства достигает центральной нервной системы ретроградным аксоплазматическим потоком. Столбнячный нейротоксин интернализуется в нервно-мышечном соединении посредством связывания белков нидоген и ретроградно транспортируется к соме в сигнальных эндосомах. Нейротропные вирусы, такие как герпесвирусы, перемещаются внутри аксонов с помощью клеточного транспортного механизма, как было показано в работе группы Элейн Беарэр. Другие инфекционные агенты также подозреваются в использовании аксонального транспорта. В настоящее время считается, что такие инфекции способствуют развитию болезни Альцгеймера и других нейродегенеративных неврологических расстройств.