
Войти
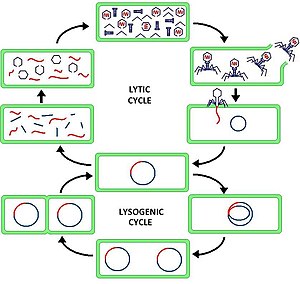 Литический цикл по сравнению с лизогенным циклом
Литический цикл по сравнению с лизогенным циклом Литический цикл ( / л ɪ т ɪ к / LIT -ik ) является одним из двух циклов вирусной репродукции ( со ссылки на бактериальные вирусы или бактериофаг ), другая являющееся лизогенной цикл. Литический цикл приводит к разрушению инфицированной клетки и ее мембраны. Бактериофаги, которые используют только литический цикл, называются вирулентными фагами (в отличие от фагов умеренного климата ).
В литическом цикле вирусная ДНК существует как отдельная свободно плавающая молекула внутри бактериальной клетки и реплицируется отдельно от бактериальной ДНК-хозяина, тогда как в лизогенном цикле вирусная ДНК находится внутри ДНК-хозяина. Это ключевое различие между литическим и лизогенным (бактерио) фаговыми циклами. Однако в обоих случаях вирус / фаг реплицируется с использованием механизма ДНК хозяина.
Литический цикл, который также называют «репродуктивным циклом» бактериофага, представляет собой шестиступенчатый цикл. Шесть стадий: прикрепление, проникновение, транскрипция, биосинтез, созревание и лизис.
Чтобы заразить клетку-хозяин, вирус должен сначала ввести в клетку свою собственную нуклеиновую кислоту через плазматическую мембрану и (если присутствует) клеточную стенку. Вирус делает это либо путем прикрепления к рецептору на поверхности клетки, либо с помощью простой механической силы. Связывание происходит из-за электростатических взаимодействий и зависит от pH и присутствия ионов. Затем вирус высвобождает свой генетический материал (одно- или двухцепочечную РНК или ДНК ) в клетку. У некоторых вирусов этот генетический материал является кольцевым и имитирует бактериальную плазмиду. На этом этапе клетка заражается и также может стать мишенью для иммунной системы. В основном этому способствуют рецепторы на поверхности клетки.
На этапах транскрипции и биосинтеза вирус захватывает механизмы репликации и трансляции клетки, используя их для создания большего количества вирусов. Нуклеиновая кислота вируса использует метаболический аппарат клетки-хозяина для производства большого количества вирусных компонентов.
В случае ДНК-вирусов ДНК транскрибируется в молекулы информационной РНК (мРНК), которые затем используются для управления рибосомами клетки. Один из первых транслируемых полипептидов разрушает ДНК хозяина. В ретровирусах (которые вводят цепь РНК) уникальный фермент, называемый обратной транскриптазой, транскрибирует вирусную РНК в ДНК, которая затем снова транскрибируется в РНК. Как только вирусная ДНК берет на себя управление, она побуждает механизм клетки-хозяина синтезировать вирусную ДНК, белок и начинает размножаться.
Биосинтез (например, Т4 ) регулируется в трех фазах продукции мРНК, за которыми следует фаза продукции белка.
Примерно через 25 минут после первоначального заражения образуется около 200 новых вирионов (вирусных телец). После созревания и накопления достаточного количества вирионов используются специализированные вирусные белки для растворения клеточной стенки. Клетка разрывается (т.е. подвергается лизису ) из-за высокого внутреннего осмотического давления (давления воды), которое больше не может сдерживаться клеточной стенкой. Это высвобождает вирионы потомства в окружающую среду, где они могут инфицировать другие клетки, и начинается еще один литический цикл. Фаг, вызывающий лизис хозяина, называется литическим или вирулентным фагом.
В геноме фага есть три класса генов, которые регулируют возникновение литического или лизогенного цикла. Первый класс - это гены непосредственного раннего развития, второй - гены отсроченного раннего периода, а третий - гены позднего периода. Следующее относится к хорошо изученному умеренному фагу лямбда E. coli.
Q-опосредованное включение поздней транскрипции начинается примерно через 6-8 мин после инфицирования, если выбран литический путь. Более 25 генов экспрессируются с одного позднего промотора, что приводит к четырем параллельным путям биосинтеза. Три пути служат для производства трех компонентов вириона: наполненной ДНК головы, хвоста и боковых волокон хвоста. Вирионы самостоятельно собираются из этих компонентов, причем первый вирион появляется примерно через 20 минут после заражения. Четвертый путь - лизис. В лямбда-5 белки участвуют в лизисе: холин и антихолин из гена S, эндолизин из гена R и белки спанина из генов Rz и Rz1. В лямбда дикого типа лизис происходит примерно через 50 мин, высвобождая примерно 100 завершенных вирионов. Время лизиса определяется белками холина и антихолина, причем последний ингибирует первый. В общем, белок холин накапливается в цитоплазматической мембране до тех пор, пока внезапно не образуются отверстия микронного размера, которые запускают лизис. Эндолизин R попадает в периплазму, где атакует пептидогликан. Белки спанина Rz и Rz1 накапливаются в цитоплазматической и внешней мембранах соответственно и образуют комплекс, охватывающий периплазму через сеть пептидогликана. Когда эндолизин разрушает пептидогликан, комплексы спанина высвобождаются и вызывают разрушение внешней мембраны. Разрушение пептидогликана эндолизином и разрушение внешней мембраны комплексом спанина оба необходимы для лизиса при лямбда-инфекциях.
Ингибирование лизиса: Т4-подобные фаги имеют два гена, rI и rIII, которые ингибируют холин Т4, если инфицированная клетка подвергается суперинфекции другим Т4 (или близкородственным) вирионом. Повторная суперинфекция может привести к тому, что инфекция Т4 продолжится без лизиса в течение нескольких часов, что приведет к накоплению вирионов до уровней, в 10 раз превышающих нормальные.