
Войти
| Повреждение нерва | |
|---|---|
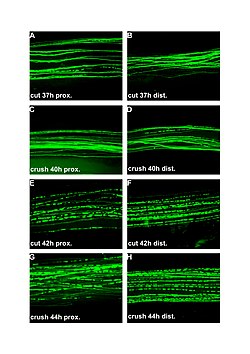 | |
| Флуоресцентные микрофотографии (100x) валлеровской дегенерации в перерезанных и раздавленных периферических нервах. Левая колонка проксимальнее травмы, правая - дистальнее. A и B: 37 часов после резки. C и D: 40 часов после увлечения. E и F: 42 часа после резки. G и H: 44 часа после травмы. | |
| Специальность | Неврология |
Валлеровская дегенерация - это активный процесс дегенерации, который возникает, когда нервное волокно разрезано или раздавлено, а часть аксона дистальнее повреждения (т.е. дальше от тела клетки нейрона ) дегенерирует. Связанный с этим процесс отмирания или ретроградной дегенерации, известный как «валлеровская дегенерация», происходит при многих нейродегенеративных заболеваниях, особенно тех, при которых нарушен транспорт аксонов, таких как БАС и болезнь Альцгеймера.. Исследования первичной культуры показывают, что неспособность доставить достаточное количество необходимого аксонального белка NMNAT2 является ключевым исходным событием.
Валлеровская дегенерация происходит после повреждение аксонов как в периферической нервной системе (ПНС), так и в центральной нервной системе (ЦНС). Это происходит в отделе аксона дистальнее места повреждения и обычно начинается в течение 24–36 часов после поражения. До дегенерации дистальный отдел аксона имеет тенденцию оставаться электрически возбудимым. После травмы аксональный скелет распадается, и аксональная мембрана разрушается. Дегенерация аксонов сопровождается деградацией миелиновой оболочки и инфильтрацией макрофагами. Макрофаги, сопровождаемые шванновскими клетками, служат для очистки остатков от дегенерации.
Шванновские клетки реагируют на потерю аксонов экструзией своих миелиновых оболочек, подавлением генов миелина, дедифференцировкой и распространение. В конце концов они выстраиваются в трубочки (ленты Бюнгнера) и экспрессируют поверхностные молекулы, направляющие регенерирующие волокна. В течение 4 дней после травмы дистальный конец части нервного волокна, проксимальной к поражению, посылает отростки к этим трубкам, и эти отростки привлекаются факторами роста, продуцируемыми шванновскими клетками в трубках. Если росток достигает трубки, он прорастает в нее и продвигается примерно на 1 мм в день, в конечном итоге достигая и повторно иннервируя ткань-мишень. Если ростки не могут добраться до трубки, например, из-за слишком широкой щели или образования рубцовой ткани, хирургическое вмешательство может помочь направить ростки в трубки. Регенерация ПНС эффективна, с почти полным восстановлением в случае поражений, которые возникают вблизи дистального окончания нервного окончания. Однако в спинном мозге выздоровление практически не наблюдается. Одно важное отличие состоит в том, что в ЦНС, включая спинной мозг, миелиновые оболочки производятся олигодендроцитами, а не шванновскими клетками.
Валлериан дегенерация названа в честь Августа Волни Уоллера. В 1850 году Уоллер экспериментировал с лягушками, перерезав им языкоглоточный и подъязычный нервы. Затем он осмотрел дистальные нервы от места повреждения, которые были отделены от своих клеточных тел в стволе мозга. Валлер описал распад миелина, который он назвал «мозговым веществом», на отдельные частицы различного размера. Дегенерирующие аксоны образовывали капли, которые можно было окрашивать, что позволяло изучать ход отдельных нервных волокон.
Хотя большинство реакций на травмы включают передачу сигналов притока кальция, способствующую повторному закрытию оторванных частей, повреждения аксонов изначально приводят к острой дегенерации аксонов (AAD), которая является быстрое разделение проксимального (части, близкой к телу клетки) и дистального конца в течение 30 минут после травмы. После разделения на обоих концах формируются дистрофические луковичные структуры, и перерезанные мембраны герметизируются. В дистальном сегменте происходит короткая латентная фаза, в течение которой он остается электрически возбудимым и структурно неповрежденным. Дегенерация сопровождается набуханием аксолеммы и, в конечном итоге, образованием бусинчатых аксональных сфероидов. Этот процесс занимает около 24 часов в ПНС и дольше в ЦНС. Сигнальные пути, ведущие к дегенерации аксолеммы, в настоящее время плохо изучены. Однако исследования показали, что этот процесс AAD не зависит от кальция.
Гранулярная дезинтеграция аксонального цитоскелета и внутренних органелл происходит после деградации аксолеммы. Ранние изменения включают накопление митохондрий в паранодальных областях в месте повреждения. Эндоплазматический ретикулум деградирует, митохондрии набухают и в конечном итоге распадаются. Происходит деполимеризация микротрубочек, за которой вскоре следует деградация нейрофиламентов и других компонентов цитоскелета. Распад зависит от протеаз убиквитин и кальпаин (вызванный притоком иона кальция), что позволяет предположить, что дегенерация аксонов является активным процессом, а не пассивным, как ранее неправильно понималось. Таким образом, аксон подвергается полной фрагментации. Скорость деградации зависит от типа травмы и также медленнее в ЦНС, чем в ПНС. Другим фактором, влияющим на скорость деградации, является диаметр аксона: более крупным аксонам требуется больше времени для деградации цитоскелета и, следовательно, требуется больше времени для дегенерации.
Миелин представляет собой фосфолипидную мембрану, которая оборачивается вокруг аксонов, обеспечивая им изоляцию. Он продуцируется клетками Шванна в ПНС и олигодендроцитами в ЦНС. Очистка от миелина - следующий шаг в дегенерации Валлера после дегенерации аксонов. Очистка от остатков миелина различна для ПНС и ЦНС. ПНС намного быстрее и эффективнее очищает миелиновые остатки по сравнению с ЦНС, и шванновские клетки являются основной причиной этого различия. Другой ключевой аспект - изменение проницаемости гемато-тканевого барьера в двух системах. В ПНС проницаемость увеличивается по всей дистальной культи, но нарушение барьера в ЦНС ограничивается только местом повреждения.
Реакция шванновских клеток на повреждение аксона быстрый. Период времени до начала дегенерации аксонов. Нейрегулины, как полагают, ответственны за быструю активацию. Они активируют рецепторы ErbB2 в микроворсинках шванновских клеток, что приводит к активации митоген-активируемой протеинкиназы (MAPK). Хотя активность MAPK наблюдается, механизм восприятия повреждений шванновскими клетками еще предстоит полностью понять. Ощущение сопровождается снижением синтеза липидов миелина и в конечном итоге прекращается в течение 48 часов. Миелиновые оболочки сначала отделяются от аксонов в области резцов Шмидта-Лантермана, а затем быстро разрушаются и укорачиваются, образуя бусообразные структуры. Шванновские клетки продолжают очищать миелиновые остатки, разрушая свой собственный миелин, фагоцитоз внеклеточный миелин и привлекают макрофаги к миелиновым остаткам для дальнейшего фагоцитоза. Однако первые несколько дней макрофаги не притягиваются к региону; следовательно, до этого момента шванновские клетки играют основную роль в очистке миелина.
Шванновские клетки, как наблюдали, рекрутируют макрофаги путем высвобождения цитокинов и хемокинов после обнаружения повреждения аксонов. Рекрутирование макрофагов помогает улучшить скорость очистки от миелинового мусора. Резидентные макрофаги, присутствующие в нервах, выделяют дополнительные хемокины и цитокины, чтобы привлечь дополнительные макрофаги. Дегенерирующий нерв также производит хемотаксические молекулы макрофагов. Еще одним источником факторов рекрутирования макрофагов является сыворотка. Задержка рекрутирования макрофагов наблюдалась у мышей с дефицитом В-клеток, лишенных сывороточных антител. Эти сигнальные молекулы вместе вызывают приток макрофагов, пик которого приходится на третью неделю после травмы. В то время как клетки Шванна опосредуют начальную стадию очистки миелинового мусора, макрофаги приходят, чтобы завершить работу. Макрофагам способствуют опсонины, которые маркируют мусор для удаления. Три основные группы, обнаруженные в сыворотке, включают комплемент, пентраксины и антитела. Однако было показано, что только комплемент помогает в фагоцитозе миелинового дебриса.
Murinson et al. (2005) наблюдали, что немиелинизированные или миелинизированные шванновские клетки при контакте с поврежденным аксоном входят в клеточный цикл, что приводит к пролиферации. Наблюдаемая продолжительность деления шванновских клеток составляла приблизительно 3 дня после повреждения. Возможные источники сигнала пролиферации приписываются рецепторам ErbB2 и рецепторам ErbB3. Эта пролиферация может еще больше повысить скорость очистки миелина и играет важную роль в регенерации аксонов, наблюдаемой в ПНС. Клетки Шванна выделяют факторы роста, которые привлекают новые отростки аксонов, растущие из проксимальной культи после полной дегенерации поврежденной дистальной культи. Это приводит к возможной реиннервации клетки или органа-мишени. Однако реиннервация не обязательно идеальна, так как возможное введение в заблуждение происходит во время реиннервации проксимальных аксонов в клетки-мишени.
По сравнению со шванновскими клетками, олигодендроцитам для выживания необходимы сигналы аксонов. На стадиях своего развития олигодендроциты, которые не могут установить контакт с аксоном и получить сигналы аксонов, подвергаются апоптозу.
. Эксперименты по Валлеровской дегенерации показали, что при повреждении олигодендроциты либо подвергаются запрограммированной клеточной гибели, либо переходят в состояние покоя. Следовательно, в отличие от клеток Шванна, олигодендроциты не могут очистить миелиновые оболочки и их остатки. В экспериментах на крысах миелиновые оболочки были обнаружены на срок до 22 месяцев. Следовательно, скорость клиренса миелиновой оболочки в ЦНС очень медленная и, возможно, может быть причиной препятствий в регенерационных возможностях аксонов ЦНС, поскольку отсутствуют факторы роста, привлекающие проксимальные аксоны. Еще одна особенность, которая в конечном итоге приводит к образованию глиального рубца. Это еще больше снижает шансы на регенерацию и реиннервацию.
Олигодендроциты не могут задействовать макрофаги для удаления мусора. Поступление макрофагов в место повреждения ЦНС происходит очень медленно. В отличие от ПНС, микроглия играет жизненно важную роль в валлеровской дегенерации ЦНС. Однако их рекрутирование происходит медленнее по сравнению с рекрутированием макрофагов в PNS примерно на 3 дня. Кроме того, микроглия может быть активирована, но гипертрофирована и не может трансформироваться в полностью фагоцитарные клетки. Те микроглии, которые действительно трансформируются, эффективно очищают от мусора. Дифференцировать фагоцитарную микроглию можно путем тестирования экспрессии главного комплекса гистосовместимости (MHC) класса I и II во время дегенерации Валлерова. Скорость выведения микроглии очень медленная по сравнению с макрофагами. Возможный источник вариаций скорости клиренса может включать в себя отсутствие активности опсонина вокруг микроглии и отсутствие повышенной проницаемости гематоэнцефалического барьера. Сниженная проницаемость может еще больше препятствовать инфильтрации макрофагов в место повреждения.
Эти данные позволяют предположить, что задержка валлеровской дегенерации в ЦНС по сравнению с ПНС вызвана не задержкой дегенерации аксонов, а скорее, из-за разницы в скорости клиренса миелина в ЦНС и ПНС.
Регенерация следует за дегенерацией. Регенерация ПНС происходит быстро, что позволяет возобновлять рост до 1 миллиметра в день. Также могут потребоваться трансплантаты для соответствующей реиннервации. Он поддерживается клетками Шванна за счет высвобождения факторов роста. Регенерация ЦНС происходит намного медленнее и почти отсутствует у большинства позвоночных. Основной причиной этого может быть задержка очистки миелиновых остатков. Миелиновые остатки, присутствующие в ЦНС или ПНС, содержат несколько ингибирующих факторов. Продолжительное присутствие миелиновых остатков в ЦНС может препятствовать регенерации. Эксперимент, проведенный на тритонах, животных, которые обладают способностью к быстрой регенерации аксонов ЦНС, показал, что валлеровская дегенерация повреждения зрительного нерва занимает в среднем от 10 до 14 дней, что также свидетельствует о том, что медленный клиренс тормозит регенерацию.
В здоровых нервах фактор роста нервов (NGF) продуцируется в очень малых количествах. Однако при повреждении экспрессия мРНК NGF увеличивается в пять-семь раз в течение 14 дней. Нервные фибробласты и шванновские клетки играют важную роль в повышении экспрессии мРНК NGF. Макрофаги также стимулируют шванновские клетки и фибробласты продуцировать NGF через интерлейкин-1, полученный из макрофагов. Другие нейротрофические молекулы, продуцируемые вместе шванновскими клетками и фибробластами, включают нейротрофический фактор головного мозга, нейротрофический фактор глиальных клеток, цилиарный нейротрофический фактор, фактор ингибирования лейкемии, инсулиноподобный фактор роста и фактор роста фибробластов. Вместе эти факторы создают благоприятную среду для роста и регенерации аксонов. Помимо факторов роста, клетки Шванна также обеспечивают структурное руководство для дальнейшего усиления регенерации. Во время фазы пролиферации шванновские клетки начинают формировать линию клеток, называемую полосами Бангнера, внутри базальной ламинарной трубки. Было обнаружено, что аксоны регенерируют в тесной связи с этими клетками. Клетки Шванна активируют выработку молекулы адгезии на клеточной поверхности ниндзюрин, дополнительно способствуя росту. Эти клеточные линии направляют регенерацию аксона в правильном направлении. Возможным источником ошибки, которая может возникнуть в результате этого, является возможное несовпадение целевых ячеек, как обсуждалось ранее.
Из-за отсутствия таких благоприятных стимулирующих факторов в ЦНС, регенерация в ЦНС задерживается.
Мыши, принадлежащие к штамму C57BL / Wld, задерживают валлеровскую дегенерацию и, таким образом, позволяют изучать роль различных типов клеток и лежащих в их основе клеточных и молекулярных процессов.. Современное понимание этого процесса стало возможным благодаря экспериментам на мышах линии Wld. Мутация впервые произошла у мышей в Harlan-Olac, лаборатории по производству животных, Соединенное Королевство. Мутация Wld представляет собой аутосомно-доминантную мутацию, встречающуюся в хромосоме 4 мыши. Мутация гена представляет собой тандемную тройную повторность 85 т.п.н., встречающуюся в природе. Мутированный участок содержит два связанных гена: никотинамидмононуклеотид аденлилтрансфераза 1 (Nmnat1) и фактор убиквитинирования e4b (Ube4b). Линкерная область, кодирующая 18 аминокислот, также является частью мутации. Было показано, что защитный эффект белка Wld обусловлен активным сайтом, синтезирующим НАД в области NMNAT1.
Хотя созданный белок локализуется в ядре и едва обнаруживается в аксонах, исследования показывают, что его защитный эффект является из-за его присутствия в аксональном и терминальном отделах. Защита, обеспечиваемая белком Wld, присуща нейронам, а не окружающим опорным клеткам, и только локально защищает аксон, что указывает на то, что внутриклеточный путь ответственен за опосредование валлеровской дегенерации.
Мутация не причиняет вреда мыши. Единственный известный эффект заключается в том, что валлеровская дегенерация задерживается в среднем до трех недель после повреждения нерва. Сначала предполагалось, что мутация Wld замедляет инфильтрацию макрофагов, но недавние исследования показывают, что мутация защищает аксоны, а не замедляет макрофаги. Процесс, с помощью которого достигается защита аксонов, плохо изучен. Однако исследования показывают, что мутация Wld приводит к повышенной активности NMNAT1, что приводит к усиленному синтезу NAD. Это, в свою очередь, активирует SIRT1-зависимый процесс в ядре, вызывая изменения в транскрипции генов. НАД сам по себе может обеспечивать дополнительную защиту аксонов за счет увеличения энергетических ресурсов аксона. Однако более поздние работы вызывают сомнения в том, что NMNAT1 или NAD могут заменять полноразмерный ген Wld. Эти авторы продемонстрировали методами in vitro и in vivo, что защитный эффект сверхэкспрессии NMNAT1 или добавления NAD не защищает аксоны от дегенерации. Однако более поздние исследования показали, что NMNAT1 является защитным в сочетании с нацеливающим на аксоны пептидом, предполагая, что ключом к защите, обеспечиваемой Wld, является комбинация активности NMNAT1 и аксональной локализации, обеспечиваемой N-концевым доменом химерного белка. 57>
Обеспеченная защита аксонов задерживает начало валлеровской дегенерации. Следовательно, активация шванновских клеток должна быть отложена, поскольку они не будут обнаруживать сигналы деградации аксонов от рецепторов ErbB2. В экспериментах на мышах с мутациями Wld инфильтрация макрофагов значительно задерживалась на срок от шести до восьми дней. Однако, как только деградация аксонов началась, дегенерация протекает нормально, и, в зависимости от нервной системы, деградация следует с вышеописанной скоростью. Возможные последствия этого позднего начала - более слабые регенеративные способности у мышей. Исследования показывают, что регенерация может быть нарушена у мышей Wld, но это, вероятно, является результатом того, что окружающая среда неблагоприятна для регенерации из-за продолжающегося существования недогенерированных дистальных волокон, тогда как обычно остатки очищаются, уступая место новому росту.
Путь валлеровской дегенерации был дополнительно прояснен открытием, что стерильный альфа- и TIR-мотив, содержащий белок 1 (SARM1), играет центральную роль в пути валлеровской дегенерации. Ген был впервые идентифицирован при скрининге мутагенеза Drosophila melanogaster, и впоследствии нокауты его гомолога у мышей показали надежную защиту перерезанных аксонов, сравнимую с таковой у Wld.
SARM1 катализирует синтез и гидролиз из циклической ADP-рибозы (cADPR) от NAD до ADP-рибозы. Активация SARM1 локально запускает быстрый коллапс уровней NAD в дистальной части поврежденного аксона, который затем подвергается дегенерации. Позднее было показано, что этот коллапс уровней NAD происходит из-за того, что TIR-домен SARM1 обладает внутренней активностью расщепления NAD. Белок SARM1 имеет четыре домена, сигнал митохондриальной локализации, аутоингибиторную N-концевую область, состоящую из мотивов броненосца / HEAT, два стерильных альфа-мотива, ответственных за мультимеризацию, и С-конец рецептора Toll / интерлейкина-1, обладающий ферментативной активностью. Активации SARM1 достаточно, чтобы снизить уровень НАД и запустить путь валлеровской дегенерации.
Активность SARM1 помогает объяснить защитную природу фактора выживания NMNAT2, как было показано ферментами NMNAT для предотвращения опосредованного SARM1 истощения НАД. Эта взаимосвязь дополнительно подтверждается тем фактом, что мыши, лишенные NMNAT2, которые обычно нежизнеспособны, полностью спасаются путем делеции SARM1, в результате чего активность NMNAT2 находится выше SARM1. Другие пути передачи сигналов, способствующие дегенерации, такие как путь киназы MAP, были связаны с активацией SARM1. Было показано, что передача сигналов MAPK способствует потере NMNAT2, тем самым способствуя активации SARM1, хотя активация SARM1 также запускает каскад киназ MAP, что указывает на существование некоторой формы петли обратной связи. Одно из объяснений защитного эффекта мутации Wld заключается в том, что область NMNAT1, которая обычно локализована в соме, заменяет лабильный фактор выживания NMNAT2 для предотвращения активации SARM1, когда N-концевой участок Ube4 белка WldS локализует его в аксон. Тот факт, что повышенная выживаемость аксонов Wld происходит из-за более медленного оборота Wld по сравнению с NMNAT2, также помогает объяснить, почему нокаут SARM1 обеспечивает более длительную защиту, поскольку SARM1 будет полностью неактивным независимо от активности ингибитора, тогда как Wld в конечном итоге будет деградировать. Возможные последствия пути SARM1 в отношении здоровья человека могут быть обнаружены на моделях животных, которые демонстрируют черепно-мозговую травму, поскольку мыши, которые содержат делеции Sarm1 в дополнение к Wld, демонстрируют уменьшение повреждения аксонов после травмы. Специфические мутации в NMNAT2 связали механизм валлеровской дегенерации с двумя неврологическими заболеваниями.
| Классификация | D |
|---|