
Войти
| Узел Ранвье | |
|---|---|
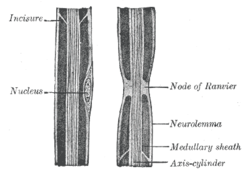 Рисунок аксона периферического нерва (обозначен как «осевой цилиндр»), показаны узел Ранвье вместе с другими особенностями Рисунок аксона периферического нерва (обозначен как «осевой цилиндр»), показаны узел Ранвье вместе с другими особенностями | |
 Узлы Ранвье Узлы Ранвье | |
| Подробности | |
| Система | Нервная система |
| Местоположение | Миелинизированный аксон нерв |
| Идентификаторы | |
| латинский | incisura myelini |
| MeSH | D011901 |
| TH | H2.00.06.2.03015 |
| Анатомические термины микроанатомии. [редактировать в Викиданных ] | |
Узлы Ранвье (, ), также известный как промежутки между миелиновой оболочкой возникают вдоль миелинизированного аксона, где аксолемма обнажена во внеклеточном пространстве. Узлы Ранвье неизолированы и сильно обогащены ионными каналами, что позволяет им участвовать в обмене ионами, необходимом для восстановления потенциала действия. Нервная проводимость в миелинизированных аксонах называется скачкообразной проводимостью (от латинского saltare «прыгать или прыгать») из-за того, как потенциал действия, кажется, «прыгает» от одного узла к другому. вдоль аксона. Это приводит к более быстрой передаче потенциала действия.
| Узел Ранвье |
|---|
 Дендрит Сома Аксон Ядро Узел. Ранвье Терминал аксона шванновская клетка Миелиновая оболочка Дендрит Сома Аксон Ядро Узел. Ранвье Терминал аксона шванновская клетка Миелиновая оболочка |
Многие аксоны позвоночных окружены миелиновой оболочкой, обеспечивающей быстрое и эффективное скачкообразное («скачущее») распространение потенциалов действия. Контакты между нейронами и глиальными клетками демонстрируют очень высокий уровень пространственной и временной организации в миелинизированных волокнах. Миелинизирующие глиальные клетки ; олигодендроциты в центральной нервной системе (ЦНС) и шванновские клетки в периферической нервной системе (ПНС), обернуты вокруг аксон, оставляя аксолемму относительно непокрытой на регулярно расположенных узлах Ранвье.
Межузловые глиальные мембраны сливаются с образованием компактного миелина, тогда как заполненные цитоплазмой паранодальные петли миелинизирующих клеток спирально обернуты вокруг аксона с обеих сторон узлов. Эта организация требует жесткого контроля развития и формирования множества специализированных зон контакта между различными участками мембраны миелинизирующих клеток. Каждый узел Ранвье окружен паранодальными областями, где спиралевидно обернутые глиальные петли прикреплены к аксональной мембране с помощью перегородочного соединения.
Сегмент между узлами Ранвье называется междоузлиями, а его самая удаленная часть, которая контактирует с параузлами, называется юкстапаранодальной областью. Узлы инкапсулированы микроворсинками, происходящими из внешней стороны мембраны шванновских клеток в ПНС, или перинодальными расширениями от астроцитов в ЦНС.
Междоузлия - это сегменты миелина, а промежутки между ними называются узлами. Размер и расстояние между междоузлиями изменяются в зависимости от диаметра волокна по криволинейной зависимости, которая оптимизирована для максимальной скорости проводимости. Размер узлов составляет от 1 до 2 мкм, тогда как междоузлия может достигать (а иногда даже больше) 1,5 мм в длину, в зависимости от диаметра аксона и типа волокна.
Структура узла и фланкирующих паранодальных областей отличается от междоузлий под компактной миелиновой оболочкой, но очень похожа в ЦНС и ПНС. Аксон подвергается воздействию внеклеточной среды в узле и сужен в своем диаметре. Уменьшение размера аксона отражает более высокую плотность упаковки нейрофиламентов в этой области, которые менее сильно фосфорилируются и транспортируются медленнее. Везикулы и другие органеллы также увеличиваются в узлах, что указывает на то, что существует узкое место для аксонального транспорта в обоих направлениях, а также для локальной аксонально-глиальной передачи сигналов.
Когда продольный разрез делается через миелинизирующую шванновскую клетку в узле, представлены три отличительных сегмента: стереотипное междоузлие, паранодальная область и узел сам. В межузловой области шванновская клетка имеет внешний воротник из цитоплазмы, компактную миелиновую оболочку и внутренний воротник из цитоплазмы, а также аксолемму. В паранодальных областях петли паранодальной цитоплазмы контактируют с утолщениями аксолеммы, образуя перегородочные соединения. В одном только узле аксолемма контактирует с несколькими микроворсинками Шванна и содержит плотную подстилку цитоскелета.
Хотя исследования переломов при замораживании показали, что узловая аксолемма как в ЦНС, так и в ПНС обогащена внутримембранными частицами (IMP) по сравнению с междоузлиями, есть некоторые структурные различия, отражающие их клеточные составляющие. В ПНС специализированные микроворсинки выступают из внешнего воротника шванновских клеток и очень близко подходят к узловой аксолемме крупных волокон. Выступы шванновских клеток перпендикулярны узлу и расходятся от центральных аксонов. Однако в ЦНС один или несколько астроцитарных процессов происходят в непосредственной близости от узлов. Исследователи заявляют, что эти процессы происходят из многофункциональных астроцитов, а не из популяции астроцитов, предназначенных для контакта с узлом. С другой стороны, в PNS базальная пластинка, которая окружает шванновские клетки, является непрерывной через узел.
Узлы Ранвье содержат Na + / K + АТФазы, обменники Na + / Ca2 + и высокую плотность потенциалзависимых каналов Na +, которые генерируют потенциалы действия. Натриевый канал состоит из порообразующей α-субъединицы и двух дополнительных β-субъединиц, которые прикрепляют канал к внеклеточным и внутриклеточным компонентам. Узлы Ранвье в центральной и периферической нервной системе в основном состоят из субъединиц αNaV1.6 и β1. Внеклеточная область субъединиц β может связываться с собой и другими белками, такими как тенасцин R и молекулы клеточной адгезии нейрофасцин и контактин. Контактин также присутствует в узлах ЦНС, и взаимодействие с этой молекулой увеличивает поверхностную экспрессию каналов Na +.
Анкирин оказался связанным с βIV спектрином, изоформой спектрина, обогащенной в узлах Ранвье и начальных сегментах аксона. Узлы PNS окружены микроворсинками шванновских клеток, которые содержат ERM и EBP50, которые могут обеспечивать связь с актиновыми микрофиламентами. Несколько белков внеклеточного матрикса обогащены в узлах Ранвье, включая тенасцин-R, Bral-1 и протеогликан NG2, а также фосфакан и versican V2. В узлах ЦНС аксональные белки также включают контактин; однако микроворсинки шванновской клетки заменены перинодальными отростками астроцитов.
Молекулярная организация узлов соответствует их специализированной функции при распространении импульсов. Уровень натриевых каналов в узле по сравнению с междоузлием предполагает, что количество IMP соответствует натриевым каналам. Калиевые каналы практически отсутствуют в узловой аксолемме, тогда как они сильно сконцентрированы в паранодальной аксолемме и мембранах шванновских клеток в узле. Точная функция калиевых каналов не совсем выяснена, но известно, что они могут способствовать быстрой реполяризации потенциалов действия или играть жизненно важную роль в буферизации ионов калия в узлах. Это сильно асимметричное распределение потенциалзависимых натриевых и калиевых каналов резко контрастирует с их диффузным распределением в немиелинизированных волокнах.
Нитевидная сеть, прилегающая к узловой мембране, содержит белки цитоскелета, называемые спектрином и анкирин. Высокая плотность анкирина в узлах может иметь функциональное значение, потому что несколько белков, которые заселяются в узлах, обладают общей способностью связываться с анкирином с чрезвычайно высоким сродством. Все эти белки, включая анкирин, обогащены начальным сегментом аксонов, что предполагает функциональную взаимосвязь. Теперь связь этих молекулярных компонентов с кластеризацией натриевых каналов в узлах все еще не известна. Хотя сообщалось, что некоторые молекулы клеточной адгезии непостоянно присутствуют в узлах; однако известно, что множество других молекул сильно заселено на глиальных мембранах паранодальных областей, где они вносят вклад в его организацию и структурную целостность.
Наблюдались сложные изменения, которые шванновская клетка претерпевает в процессе миелинизации периферических нервных волокон и изучается многими. Начальная оболочка аксона происходит без прерывания на всем протяжении шванновской клетки. Последовательность этого процесса заключается в свертывании поверхности клетки Шванна таким образом, что образуется двойная мембрана на противоположных гранях свернутой поверхности клетки Шванна. Эта мембрана растягивается и спирально оборачивается снова и снова по мере продолжения свертывания поверхности шванновской клетки. В результате легко установить увеличение толщины расширения миелиновой оболочки миелин в ее поперечном сечении. Также очевидно, что каждый из последовательных витков спирали увеличивается в размере по длине аксона по мере увеличения числа витков. Однако неясно, может ли увеличение длины миелиновой оболочки быть объяснено исключительно увеличением длины аксона, покрываемого каждым последующим витком спирали, как объяснялось ранее. На стыке двух шванновских клеток вдоль аксона направления ламеллярного выступа миелиновых окончаний имеют противоположное значение. Это соединение, смежное с шванновскими ячейками, составляет область, обозначенную как узел Ранвье.
Исследователи доказывают, что в развивающейся ЦНС, Nav1.2 изначально экспрессируется во всех формирующих узлах Ранвье. После созревания узловой Nav1.3 подавляется и заменяется Nav1.6. Naz1.2 также экспрессируется во время формирования узла PNS, что свидетельствует о том, что переключение подтипов Nav-каналов является общим явлением в CNS и PNS. В этом же исследовании было показано, что Nav1.6 и Nav1.2 совместно локализуются во многих узлах Ранвье во время ранней миелинизации. Это также привело к предположению, что ранние кластеры каналов Nav1.2 и Nav1.6 должны впоследствии стать узлами Ранвье. Нейрофасцин также считается одним из первых белков, которые накапливаются во вновь формирующихся узлах Ранвье. Также обнаружено, что они обеспечивают сайт зародышеобразования для прикрепления G анкирина, каналов Nav и других белков. Недавняя идентификация глиомедина шванновских клеток микроворсинок в качестве вероятного партнера по связыванию аксонального нейрофасцина дает существенные доказательства важности этого белка в рекрутировании Nav-каналов в узлы Ранвье. Кроме того, Lambert et al. и Eshed et al. также указывает на то, что нейрофасцин накапливается до каналов Nav и, вероятно, играет решающую роль в самых ранних событиях, связанных с образованием узла Ранвье. Таким образом, несколько механизмов могут существовать и работать синергетически для облегчения кластеризации каналов навигации в узлах Ранвье.
Первым событием, по-видимому, является накопление молекул клеточной адгезии, таких как NF186 или NrCAM. Внутриклеточные области этих молекул клеточной адгезии взаимодействуют с анкирином G, который служит якорем для натриевых каналов. В то же время периаксональное расширение глиальной клетки оборачивается вокруг аксона, давая начало паранодальным областям. Это движение вдоль аксона вносит значительный вклад в общее формирование узлов Ранвье, позволяя геминодам, сформированным на краях соседних глиальных клеток, сливаться в полные узлы. Септатоподобные соединения образуются в паранодальных узлах с обогащением NF155 в глиальных параноузловых петлях. Сразу после ранней дифференциации узловых и паранодальных областей калиевые каналы, Caspr2 и TAG1 накапливаются в прилегающих и паранодальных областях. Это накопление непосредственно совпадает с образованием компактного миелина. В зрелых узловых областях взаимодействия с внутриклеточными белками кажутся жизненно важными для стабильности всех узловых областей. В ЦНС олигодендроциты не имеют микроворсинок, но, по-видимому, способны инициировать кластеризацию некоторых аксональных белков через секретируемые факторы. Комбинированные эффекты таких факторов с последующими движениями, генерируемыми обертыванием периаксонального удлинения олигодендроцита, могут объяснить организацию узлов ЦНС Ранвье.
потенциал действия - это всплеск как положительного, так и отрицательного ионного разряда, который проходит вдоль мембраны клетки. Создание и проведение потенциалов действия представляет собой фундаментальное средство коммуникации в нервной системе. Потенциалы действия представляют собой быстрое изменение напряжения на плазматической мембране аксонов. Эти быстрые изменения опосредуются потенциалозависимыми ионными каналами, обнаруженными в плазматической мембране. Потенциал действия перемещается от одного места в клетке к другому, но поток ионов через мембрану происходит только в узлах Ранвье. В результате сигнал потенциала действия скачет по аксону от узла к узлу, а не распространяется плавно, как это происходит в аксонах, лишенных миелиновой оболочки. Кластеризация потенциалзависимых каналов ионов натрия и калия в узлах допускает такое поведение.
Поскольку аксон может быть немиелинизированным или миелинизированным, потенциал действия может перемещаться вниз по аксону двумя способами. Эти методы называются непрерывной проводимостью для немиелинизированных аксонов и скачкообразной проводимостью для миелинизированных аксонов. Сальтаторная проводимость определяется как потенциал действия, перемещающийся дискретными скачками вниз по миелинизированному аксону.
Этот процесс описывается как заряд , пассивно распространяющийся на следующий узел Ранвье, чтобы деполяризовать его до порогового значения, которое затем запускает потенциал действия в этой области, который затем будет пассивно распространяться на следующий узел. узел и так далее.
Сальтирующая проводимость обеспечивает одно преимущество перед проводимостью, которая происходит вдоль аксона без миелиновых оболочек. Дело в том, что повышенная скорость, обеспечиваемая этим режимом проведения, обеспечивает более быстрое взаимодействие между нейронами. С другой стороны, в зависимости от средней частоты возбуждения нейрона, расчеты показывают, что энергетические затраты на поддержание потенциала покоя олигодендроцитов могут перевесить экономию энергии за счет потенциалов действия. Таким образом, миелинизация аксонов не обязательно экономит энергию.
Митохондрии и другие мембранные органеллы обычно обогащены в области PNP периферических миелинизированных аксонов, особенно аксонов большого калибра. Фактическая физиологическая роль этого накопления и факторы, которые его регулируют, не изучены; однако известно, что митохондрии обычно присутствуют в областях клетки, которые проявляют высокую потребность в энергии. В этих же областях они также считаются содержащими конусы роста, синаптические окончания и сайты инициации и регенерации потенциала действия, такие как узлы Ранвье. В синаптических окончаниях митохондрии производят АТФ, необходимый для мобилизации пузырьков для нейротрансмиссии. В узлах Ранвье митохондрии играют важную роль в проведении импульсов, вырабатывая АТФ, который необходим для поддержания активности энергоемких ионных насосов. В подтверждение этого факта, в аксоплазме PNP больших периферических аксонов присутствует примерно в пять раз больше митохондрий, чем в соответствующих межузловых областях этих волокон.
Сальтаторная проводимость в миелинизированных аксонах требует организации узлов Ранвье, тогда как потенциал-управляемые натриевые каналы сильно заселены. Исследования показывают, что αII-Spectrin, компонент цитоскелета, обогащается в узлах и параузлах на ранних стадиях, и по мере созревания узлов экспрессия этой молекулы исчезает. Также доказано, что αII-Spectrin в аксональном цитоскелете абсолютно необходим для стабилизации кластеров натриевых каналов и организации зрелого узла Ранвье.
Ранее было показано, что OMgp (олигодендроцитарный миелиновый гликопротеин) кластеризуется в узлах Ранвье и может регулировать параночечную архитектуру, длину узла и разрастание аксонов в узлах. Однако последующее исследование показало, что антитело, которое ранее использовалось для идентификации OMgp в узлах, перекрестно реагирует с другим обогащенным узлами компонентом versican V2 и что OMgp не требуется для целостности узлов и параузлов, что является аргументом против ранее сообщалось о локализации и предполагаемых функциях OMgp в узлах.
Белки в этих возбудимых доменах нейрона при повреждении могут приводить к когнитивным нарушениям и различным невропатическим заболеваниям.
 Луи Антуан Ранвье (1835–1922)
Луи Антуан Ранвье (1835–1922) Миелиновая оболочка длинных нервов была обнаружена и названа немецким патологическим анатомом Рудольфом Вирховым в 1854 г. Французский патолог и анатом Луи-Антуан Ранвье позже обнаружил узлы или щели в миелиновой оболочке, которые теперь носят его имя. Ранвье родился в Лионе и был одним из самых выдающихся гистологов конца XIX века. Ранвье оставил патологические исследования в 1867 году и стал помощником физиолога Клода Бернара. Он был председателем кафедры общей анатомии в Collège de France в 1875 году.
Его усовершенствованные гистологические методы и его работа как с поврежденными, так и с нормальными нервными волокнами стали мировыми. известный. Его наблюдения за волокнистыми узлами и дегенерацией и регенерацией разрезанных волокон оказали большое влияние на парижскую неврологию в Сальпетриере. Вскоре после этого он обнаружил разрывы в оболочках нервных волокон, которые позже были названы Узлами Ранвье. Это открытие позже привело Ранвье к тщательному гистологическому исследованию миелиновых оболочек и шванновских клеток.

Полная диаграмма нейронных клеток

Медуллированные нервные волокна, окрашенные нитратом серебра