
Войти
| АДФ / АТФ-транслоказы | |||||||||
|---|---|---|---|---|---|---|---|---|---|
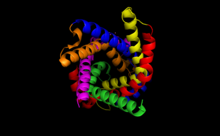 Цитоплазматический вид связывающего кармана АТФ-АДФ-транслоказы 1, PDB : 1OKC.) Цитоплазматический вид связывающего кармана АТФ-АДФ-транслоказы 1, PDB : 1OKC.) | |||||||||
| Идентификаторы | |||||||||
| Символ | Aden_trnslctor | ||||||||
| Pfam | PF00153 | ||||||||
| InterPro | IPR002113 | ||||||||
| TCDB | 2.A.29.1.2 | ||||||||
| суперсемейство OPM | 21 | ||||||||
| белок OPM | 2c3e | ||||||||
| |||||||||
| семейство 25 переносчиков растворенных веществ (митохондриальный переносчик; транслокатор адениновых нуклеотидов), член 4 | |
|---|---|
| Идентификаторы | |
| Символ | SLC25A4 |
| Альт. символы | PEO3, PEO2, ANT1 |
| Ген NCBI | 291 |
| HGNC | 10990 |
| OMIM | 103220 |
| RefSeq | NM_001151 |
| UniProt | P12235 |
| Прочие данные | |
| Locus | Chr. 4 q35 |
| семейство 25 носителей растворенного вещества (митохондриальный носитель; транслокатор адениновых нуклеотидов), член 5 | |
|---|---|
| Идентификаторы | |
| Символ | SLC25A5 |
| Альт. символы | ANT2 |
| ген NCBI | 292 |
| HGNC | 10991 |
| OMIM | 300150 |
| RefSeq | NM_001152 |
| UniProt | P05141 |
| Прочие данные | |
| Locus | Chr. X q24-q26 |
| семейство 25 переносчиков растворенного вещества (митохондриальный переносчик; транслокатор адениновых нуклеотидов), член 6 | |
|---|---|
| Идентификаторы | |
| Символ | SLC25A6 |
| Альт. символы | ANT3 |
| ген NCBI | 293 |
| HGNC | 10992 |
| OMIM | 403000 |
| RefSeq | NM_001636 |
| UniProt | P12236 |
| Прочие данные | |
| Locus | Chr. Y p |
транслокатор адениновых нуклеотидов (ANT ), также известный как транслоказа АДФ / АТФ (ANT ), АДФ / АТФ белок-носитель (AAC ) или митохондриальный переносчик АДФ / АТФ, обменивает свободный АТФ на свободный АДФ через внутреннюю митохондриальную мембрану. АНТ является наиболее распространенным белком внутренней митохондриальной мембраны и принадлежит к семейству митохондриальных переносчиков.
Свободный АДФ транспортируется из цитоплазмы в матрикс митохондрий, а АТФ образующийся в результате окислительного фосфорилирования, переносится из митохондриального матрикса в цитоплазму, обеспечивая таким образом клетки своей основной энергетической валютой. Транслоказы АДФ / АТФ являются эксклюзивными для эукариот и, как полагают, эволюционировали во время эукариогенеза. Клетки человека экспрессируют четыре транслоказы АДФ / АТФ: SLC25A4, SLC25A5, SLC25A6 и SLC25A31, которые составляют более 10% белка. во внутренней митохондриальной мембране. Эти белки классифицируются в надсемействе митохондриальный носитель.
У людей существуют три параологичных ANT изоформы :
ANT имеет долгое время считалось, что он функционирует как гомодимер, но эта концепция была поставлена под сомнение из-за проекционной структуры дрожжевого Aac3p, решенной с помощью электронной кристаллографии, которая показала, что белок является трехкратно симметричным и мономерным, с путем транслокации субстрата через центр.Атомная структура бычьего ANT подтвердила это представление и предоставила первую структурную складку митохондриального носителя. Дальнейшие исследования показали, что ANT является мономером в детергентах и функционирует как мономер в митохондриальных мембранах.
ADP / АТФ-транслоказа 1 является основным AAC в клетке человека s и архетипический белок этого семейства. Он имеет массу примерно 30 кДа и состоит из 297 остатков. Он образует шесть трансмембранных α-спиралей, которые образуют бочку, которая приводит к глубокому конусообразному углублению, доступному снаружи, где связывается субстрат. Связывающий карман, консервативный во всех изоформ, в основном состоит из основных остатков, которые позволяют прочно связываться с АТФ или АДФ, и имеет максимальный диаметр 20 Å и глубину 30 Å. Действительно, было показано, что аргинин, остатки 96, 204, 252, 253 и 294, а также лизин 38 важны для активности транспортера.
АДФ / АТФ-транслоказа транспортирует АТФ, синтезированный в результате окислительного фосфорилирования, в цитоплазму, где его можно использовать в качестве основной энергетической валюты клетки для обеспечения термодинамически неблагоприятных реакций. После последующего гидролиза АТФ до АДФ, АДФ транспортируется обратно в митохондриальный матрикс, где он может быть повторно фосфорилирован до АТФ. Поскольку человек обычно ежедневно обменивает эквивалент своей собственной массы АТФ, транслоказа АДФ / АТФ является важным белком-переносчиком с основными метаболическими нарушениями.
ANT переносит свободный, т.е. депротонированные, несвязанные с магнием, несвязанные с кальцием формы АДФ и АТФ в соотношении 1: 1. Транспорт полностью обратим, и его направленность определяется концентрацией его субстратов (АДФ и АТФ внутри и вне митохондрий), хелаторов адениновых нуклеотидов и потенциала митохондриальной мембраны. Взаимосвязь этих параметров может быть выражена уравнением, решающим «потенциал реверсии ANT» (Erev_ANT), значение потенциала митохондриальной мембраны, при котором ANT не осуществляет чистый транспорт адениновых нуклеотидов. ANT и АТФ-синтаза F0-F1 не обязательно находится в направленной синхронности.
Помимо обмена АДФ и АТФ через внутреннюю митохондриальную мембрану, ANT также проявляет внутреннюю активность разобщения
ANT является важным модулирующим и возможным структурным компонентом поры перехода митохондриальной проницаемости, канала, участвующего в различных патологиях, функция которых все еще остается неуловимой. Karch et al. Предлагают «многопорную модель», в которой ANT является по крайней мере одним из молекулярные компоненты поры.
В нормальных условиях АТФ и АДФ не могут пересекать внутреннюю митохондриальную мембрану из-за их высокого отрицательного заряда, но АДФ / АТФ-транслоказа антипортер, связывает транспорт двух молекул. Депрессия в транслоказе АДФ / АТФ альтернативно обращена к матриксу и цитоплазматической стороне мембраны. АДФ в межмембранном пространстве, исходящий из цитоплазмы, связывает транслоказу и вызывает ее выворот, что приводит к высвобождению АДФ в матрикс. Связывание АТФ с матрицей вызывает эверсию и приводит к высвобождению АТФ в межмембранное пространство, впоследствии диффундирующего в цитоплазму, и одновременно возвращает транслоказу к ее исходной конформации. АТФ и АДФ - единственные природные нуклеотиды, распознаваемые транслоказой.
Сетевой процесс обозначается:
Обмен АДФ / АТФ является энергетически дорогостоящим: около 25% энергии выделяется в результате переноса электрона посредством аэробного дыхания, или один ион водорода, расходуется для восстановления мембранного потенциала, который задействуется АДФ / АТФ-транслоказой.
Транслокатор циклически переключается между двумя состояниями, называется цитоплазматическим и матричным состоянием, открываясь этим компартментам поочередно. Существуют доступные структуры, которые показывают, что транслокатор заблокирован в цитоплазматическом состоянии ингибитором карбоксиатрактилозидом или в матричном состоянии ингибитором бонгкрековой кислотой.
Редко, но тяжелые заболевания, такие как митохондриальные миопатии, связаны с дисфункциональной транслоказой АДФ / АТФ человека. Митохондриальные миопатии (ММ) относятся к группе клинически и биохимически гетерогенных заболеваний, которые имеют общие черты основных митохондриальных структурных аномалий в скелетных мышцах. Основным морфологическим признаком ММ являются рваные красные волокна, содержащие периферические и интермиофибриллярные скопления аномальных митохондрий. В частности, аутосомно-доминантная прогрессирующая внешняя офтальмоплегия (adPEO) является распространенным заболеванием, связанным с дисфункциональной транслоказой ADP / ATP, и может вызывать паралич мышц, ответственных за движения глаз. Общие симптомы не ограничиваются глазами и могут включать непереносимость физических упражнений, мышечную слабость, нарушение слуха и многое другое. adPEO демонстрирует образцы менделевского наследования, но характеризуется крупномасштабными делециями митохондриальной ДНК (мтДНК). мтДНК содержит несколько интронов или некодирующих областей ДНК, что увеличивает вероятность вредных мутаций. Таким образом, любая модификация мтДНК АДФ / АТФ транслоказы может привести к нарушению функции переносчика, особенно к остаткам, вовлеченным в связывающий карман, что может поставить под угрозу эффективность транслоказы. ММ обычно ассоциируется с дисфункциональной транслоказой АДФ / АТФ, но ММ может индуцироваться через множество различных митохондриальных аномалий.
 Бонкрекиновая кислота
Бонкрекиновая кислота АДФ / АТФ-транслоказа очень специфично ингибируется двумя семействами соединений. Первое семейство, которое включает атрактилозид (ATR) и карбоксиатрактилозид (CATR), связывается с транслоказой ADP / ATP с цитоплазматической стороны, запирая ее в открытой конформации цитоплазматической стороны. Напротив, второе семейство, которое включает бонгкрековую кислоту (BA) и изобонгкрековую кислоту (isoBA), связывает транслоказу из матрикса, запирая ее в открытой конформации на стороне матрикса. Отрицательно заряженные группы ингибиторов прочно связываются с положительно заряженными остатками глубоко внутри связывающего кармана. Высокое сродство (Kd в наномолярном диапазоне) делает каждый ингибитор смертельным ядом, затрудняя клеточное дыхание / передачу энергии остальной части клетки. Существуют доступные структуры, которые показывают, что транслокатор заблокирован в цитоплазматическом состоянии ингибитором карбоксиатрактилоидом или в матричном состоянии ингибитором бонгкрековой кислотой.
В 1955 г., Siekevitz и Potter продемонстрировали, что нуклеотиды аденина были распределены в клетках в двух пулах, расположенных в митохондриальном и цитозольном компартментах. Вскоре после этого Прессман предположил, что два пула могут обмениваться нуклеотидами. Однако существование переносчика АДФ / АТФ не предполагалось до 1964 г., когда Bruni et al. открыли ингибирующее действие атрактилоида на систему передачи энергии (окислительное фосфорилирование) и сайты связывания АДФ в митохондриях печени крыс. Вскоре после этого было проведено огромное количество исследований для доказательства существования и выяснения связи между транслоказой АДФ / АТФ и транспортом энергии. кДНК транслоказы АДФ / АТФ была секвенирована для крупного рогатого скота в 1982 г. и видов дрожжей Saccharomyces cerevisiae в 1986 году, прежде чем, наконец, Battini et al. секвенировал клон кДНК человеческого транспортера в 1989 году. Гомология в кодирующих последовательностях между АДФ / АТФ транслоказой человека и дрожжей составила 47%, в то время как бычьи и человеческие последовательности значительно расширились до 266 из 297 остатков, или 89,6 %. В обоих случаях наиболее консервативные остатки находятся в кармане связывания субстрата АДФ / АТФ.