
Войти
KcsA (Kcканал s treptomyces A) является прокариотическим калиевый канал из почвенных бактерий Streptomyces lividans, который широко изучался в исследовании ионного канала. Активированный белок pH обладает двумя трансмембранными сегментами и высокоселективной областью поры, ответственной за выход и перемещение ионов К из клетки. Аминокислотная последовательность, обнаруженная в фильтре селективности KcsA, высококонсервативна как для прокариотических, так и для эукариотических K-каналов напряжения; В результате исследования KcsA предоставили важную структурную и механистическую информацию о молекулярных основах отбора и проводимости ионов K. Как один из наиболее изученных ионных каналов и по сей день, KcsA является шаблоном для исследования функции K-канала, и его выясненная структура лежит в основе компьютерного моделирования динамики каналов как для прокариот, так и для эукариот.
| Калиевый канал KcsA | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Четыре субъединицы, образующие канал, нарисованы разными цветами. Они окружают центральную пору, защищенную фильтром селективности, состоящим из P-петель каждой из субъединиц. Синие и красные точки указывают границы липидного бислоя. Четыре субъединицы, образующие канал, нарисованы разными цветами. Они окружают центральную пору, защищенную фильтром селективности, состоящим из P-петель каждой из субъединиц. Синие и красные точки указывают границы липидного бислоя. | |||||||||
| Идентификаторы | |||||||||
| Символ | KcsA | ||||||||
| Pfam | PF07885 | ||||||||
| InterPro | IPR013099 | ||||||||
| SCOPe | 1bl8 / SUPFAM | ||||||||
| OPM суперсемейство | 8 | ||||||||
| OPM белок | 1r3j | ||||||||
| |||||||||
KcsA был первым каналом для ионов калия, который был охарактеризован с помощью рентгеновской кристаллографии Родериком Маккинноном и его коллегами в 1998 году. В годы, предшествовавшие этому, исследования структуры К-каналов были сосредоточены на использовании малых токсинов связывание для выявления местоположения поры и фильтра селективности среди остатков канала. Группа Маккиннона теоретизировала тетрамерное расположение трансмембранных сегментов и даже предположила наличие порообразующих «петель» в фильтрующей области, состоящей из коротких сегментов аминокислот, которые взаимодействуют с Ионы K проходят через канал Обнаружение сильной последовательности гомологии между KcsA и другими каналами семейства Kv, включая белок Shaker, привлекло внимание научного сообщества. особенно после того, как последовательность сигнатуры K-канала стала появляться в других прокариотических генах. Простота двух трансмембранных спиралей в KcsA, в отличие от шести во многих эукариотических ионных каналах, также предоставила метод для понимания механизмов проведения K-каналов на более рудиментарном уровне, тем самым обеспечивая даже большой импульс. для изучения KcsA.
Кристаллическая структура KcsA была решена группой MacKinnon в 1998 году после открытия, что удаление С-конца цитоплазматического домена нативного белка (остатки 126-158) увеличивает стабильность кристаллизованные образцы. Была получена модель KcsA с разрешением 3,2А, которая подтвердила тетрамерное расположение белка вокруг центральной поры, причем одна спираль каждой субъединицы обращена к внутренней оси, а другая - наружу. Три года спустя Мораис-Кабрал и Чжоу создали модель с более высоким разрешением после присоединения моноклональных фрагментов Fab к кристаллам KcsA для дальнейшей стабилизации канала. В начале 2000-х годов на основе энергетических и электростатических расчетов, сделанных для моделирования области поры, появилось свидетельство заполнения фильтра селективности двумя атомами K во время процесса переноса. Продолжение исследований различных открытых и закрытых, неактивных и активных конформаций KcsA с помощью других методов визуализации, таких как ssNMR и EPR, с тех пор предоставило еще большее понимание структуры канала и сил, ограничивающих переключиться с инактивации канала на проводимость.
В 2007 году Riek et. Al. показали, что открытие канала, которое возникает в результате титрования ионного канала с pH 7 до pH 4, соответствует конформационным изменениям в двух областях: переходу в ионообменное состояние фильтра селективности и открытию расположение TM2 на C-конце. Эта модель объясняет способность KcsA одновременно отбирать ионы K, одновременно контролируя электрическую проводимость. В 2011 г. кристаллическая структура полноразмерного KcsA была разрешена, чтобы выявить, что препятствие со стороны ранее усеченных остатков позволяет только прямое расширение области межклеточного прохождения ионов белка. Это исследование позволяет более подробно изучить движение отдельных участков канала при ионной проводимости. В настоящее время исследования KcsA сосредоточены на использовании прокариотического канала в качестве модели для динамики каналов более крупных эукариотических K-каналов, включая hERG.
 Кристаллическая структура KcsA. Здесь показаны только два из четырех субъединиц. Белок показан зеленым цветом, карбонильные группы основной цепи (кислород = красный, углерод = зеленый) и ионы калия (занимающие позиции S2 и S4) и атомы кислорода молекул воды (S1 и S3) представляют собой пурпурные и красные сферы соответственно.
Кристаллическая структура KcsA. Здесь показаны только два из четырех субъединиц. Белок показан зеленым цветом, карбонильные группы основной цепи (кислород = красный, углерод = зеленый) и ионы калия (занимающие позиции S2 и S4) и атомы кислорода молекул воды (S1 и S3) представляют собой пурпурные и красные сферы соответственно. Структура KcsA представляет собой перевернутый конус, с центральной порой, идущей вниз по центру, состоящей из двух трансмембранных спиралей (внешней спирали M1 и внутренней спирали M2), которые охватывают липидный бислой. Сам канал представляет собой тетрамер , состоящий из четырех идентичных однодоменных субъединиц (каждая с двумя α-спиралями), расположенных так, что одна спираль M2 обращена к центральной поре, а другая спираль M1 обращена к липидная мембрана. Внутренние спирали наклонены примерно на 25 ° по отношению к липидной мембране и слегка изогнуты, открываясь, обращаясь к внешней стороне клетки, как цветок. Эти две ТМ-спирали связаны возвратной петлей, симметрично распределенной вокруг общей оси, соответствующей центральной поре. Область пор охватывает примерно 30 аминокислотных остатков и может быть разделена на три части: фильтр селективности около внеклеточной стороны, расширенная заполненная водой полость в центре и сформированный закрытый затвор около цитоплазматической стороны. четырьмя упакованными спиралями М2. Обнаружено, что эта архитектура высококонсервативна в семействе калиевых каналов как у эукариот, так и у прокариот.
Общая длина поры составляет 45 Å, а ее диаметр значительно варьируется в отдельных областях внутреннего туннеля. Двигаясь от внутриклеточной области наружу (снизу вверх на рисунке), пор начинается с области затвора, образованной спиралями M2 диаметром 18 Å, а затем открывается в широкую полость. (∼10 Å в поперечнике) около середины мембраны. В этих областях ионы K находятся в контакте с окружающими молекулами воды, но когда они входят в канал из фильтра селективности вверху, полость настолько узкая, что ионы K должны отводить любую гидратирующую воду, чтобы попасть в ячейку. Что касается аминокислотного состава остатков, выстилающих поры в KcsA, боковые цепи, выстилающие внутреннюю пору и полость, преимущественно гидрофобны, но в пределах фильтра селективности полярны присутствуют аминокислоты, которые контактируют с дегидратированными ионами K.
Более широкий конец конуса соответствует внеклеточному устью канала, состоящему из спиралей пор, плюс фильтр селективности который образован последовательностью TVGYG (треонин, валин, глицин, тирозин, глицин), характерной для калиевых каналов. В этой области координация между аминокислотами TVGYG и поступающими ионами K позволяет проводить ионы через канал. Фильтр селективности KcsA содержит четыре сайта связывания ионов, хотя предполагается, что только два из этих четырех положений заняты одновременно. Фильтр селективности имеет диаметр около 3 Å. хотя молекулярная динамика моделирования предполагает, что фильтр гибкий. Присутствие TVGYG в фильтрующей области KcsA сохраняется даже в более сложных эукариотических каналах, что делает KcsA оптимальной системой для изучения проводимости K-канала у разных видов.
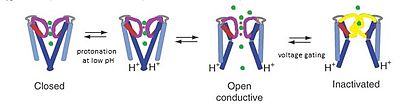 KcsA переходит из закрытой в открытую конформацию при протонировании спирали M2 при низком pH. Стробирование напряжения приводит к разрушению фильтра селективности и последующей деактивации. Изображение адаптировано из Thompson et al. 2008.
KcsA переходит из закрытой в открытую конформацию при протонировании спирали M2 при низком pH. Стробирование напряжения приводит к разрушению фильтра селективности и последующей деактивации. Изображение адаптировано из Thompson et al. 2008. Канал KcsA считается моделью канала, потому что структура KcsA обеспечивает основу для понимания проводимости K-канала, которая состоит из трех частей: Калий селективность, блокирование канала с помощью чувствительности к pH и инактивация канала с ограничением по напряжению. Проникновение ионов K происходит в верхней области селективного фильтра поры, в то время как регулирование pH возникает из-за протонирования трансмембранных спиралей в точке конец поры. При низком pH спираль M2 протонируется, смещая ионный канал из закрытой в открытую конформацию. Считается, что когда ионы проходят через канал, механизмы стробирования напряжения вызывают взаимодействия между Glu71 и Asp80 в фильтре селективности, которые дестабилизируют проводящую конформацию и облегчают переход в долгоживущее непроводящее состояние, которое напоминает инактивацию C-типа потенциалозависимые каналы.
В непроводящей конформации KcsA при pH 7, K тесно связан с координирующими атомами кислорода фильтра селективности, и четыре спирали TM2 сходятся около цитоплазматической переход, чтобы заблокировать прохождение любых ионов калия. Однако при pH 4 KcsA претерпевает конформационные обмены в миллисекундном масштабе времени, фильтруя проницаемые и непроницаемые состояния, а также между открытой и закрытой конформациями спиралей M2. Хотя эти отчетливые конформационные изменения происходят в отдельных областях канала, молекулярное поведение каждой области связано как электростатическими взаимодействиями, так и аллостерией. Динамика этой обменной стереохимической конфигурации в фильтре обеспечивает физическую основу для одновременной K-проводимости и стробирования.
Последовательность TVGYG особенно важна для поддержания калиевой специфичности KcsA. Глицины в этой последовательности фильтров селективности имеют двугранные углы, которые позволяют карбонильным атомам кислорода в белковой основе фильтра указывать в одном направлении, к ионам вдоль поры. Глицины и треонин координируются с ионом K, в то время как боковые цепи валина и тирозина направляются в ядро белка, чтобы наложить геометрические ограничения на фильтр. В результате тетрамер KcsA содержит четыре равных друг другу сайта связывания K, каждая сторона которых состоит из клетки, образованной восемью атомами кислорода, которые находятся в вершинах куба. Атомы кислорода, которые окружают ионы K в фильтре, расположены подобно молекулам воды, которые окружают гидратированные ионы K в полости канала; это говорит о том, что центры координации и связывания кислорода в фильтре селективности оплачивают энергетические затраты на дегидратацию калия. Поскольку ион Na + слишком мал для этих сайтов связывания размера K, энергия дегидратации не компенсируется, и, таким образом, фильтр осуществляет селекцию против других посторонних ионов. Кроме того, канал KcsA заблокирован ионами Cs, а для стробирования требуется присутствие ионов Mg.
PH-зависимая проводимость KcsA указывает на то, что открытие ионного канала происходит, когда белок подвергается воздействию более кислой среды. Исследования ЯМР, проведенные группой Riek, показывают, что чувствительность к pH проявляется как в C-концевой области TM2 белка, так и в остатках Tyr78 и Gly79 в фильтре селективности. Есть основания полагать, что основной датчик pH находится в цитоплазматическом домене. Замена отрицательно заряженных аминокислот на нейтральные сделала канал KcsA нечувствительным к pH, даже несмотря на отсутствие аминокислотных изменений в трансмембранной области. Кроме того, при pH от 6 до 7 гистидин является одной из немногих титруемых боковых цепей гистидинов; они отсутствуют в трансмембранных и внеклеточных сегментах TM2, но присутствуют на C-конце KcsA. Это подчеркивает возможный механизм медленного открытия KcsA, который особенно чувствителен к pH, особенно потому, что конформационное распространение сигнала открытия канала от C-конца к фильтру селективности может быть важным для координации структурных изменений, необходимых для проводимости вдоль всей поры..
Исследования ЯМР также предполагают, что сложная сеть водородных связей между Tyr78, Gly79, Glu71 и Asp80 существует в области фильтра KcsA и, кроме того, действует как pH-чувствительный триггер для проводимости. Мутация ключевых остатков в регионе, включая E71A, приводит к большим затратам энергии в 4 ккал моль, что эквивалентно потере водородной связи между Glu71 и Tyr78 и опосредованной водой водородной связи между Glu71 и Asp80 в KcsA (E71A). Эти исследования дополнительно подчеркивают роль гейтирования pH в функции канала KcsA.
В 2006 году группа Perozo предложила механистическое объяснение влияния полей напряжения на стробирование KcsA. После добавления в канал деполяризующего тока происходит переориентация Glu71 по направлению к внутриклеточной поре, тем самым разрушая карбоксилкарбоксилатную пару Glu71-Asp80, которая изначально стабилизирует фильтр селективности. Сжатие области фильтра предотвращает вход в неактивное состояние или облегчает выход из него. Glu71, ключевая часть последовательности сигнатуры фильтра селективности, которая сохраняется среди ионных каналов K, играет ключевую роль в стробировании, поскольку его способность переориентировать себя в направлении трансмембранного поля напряжения способна дать объяснение событий стробирования напряжения в KcsA. Ориентация аминокислот в фильтрующей области может играть важную физиологическую роль в модуляции потоков калия у эукариот и прокариот в стационарных условиях.
Точная Механизм селективности калиевых каналов продолжает изучаться и обсуждаться, и для описания различных аспектов селективности используется множество моделей. К KcsA были применены модели, объясняющие избирательность на основе концепции напряженности поля, разработанной Джорджем Эйзенманом на основе закона Кулона. Альтернативное объяснение избирательности KcsA основано на модели плотного прилегания (также известной как модель плотного прилегания), разработанной Benzanilla и Armstrong. Карбонильные атомы кислорода основной цепи, составляющие фильтр селективности, удерживаются в точном положении, которое позволяет им заменять молекулы воды в гидратированной оболочке калия ион, но они слишком далеки от иона натрия. Дальнейшая работа была посвящена изучению термодинамических различий в связывании ионов, топологических соображениях и количестве участков непрерывного связывания ионов.
Кроме того, основное ограничение исследования кристаллической структуры и моделирования еще предстоит обсуждалось: наиболее разрешенная и наиболее применяемая кристаллическая структура KcsA, по-видимому, представляет собой «закрытую» форму канала. Это разумно, поскольку закрытое состояние канала благоприятно при нейтральном pH, при котором кристаллическая структура была определена с помощью рентгеновской кристаллографии. Однако динамическое поведение KcsA затрудняет анализ канала, поскольку кристаллическая структура неизбежно обеспечивает статическое, усредненное по пространству и времени изображение канала. Чтобы преодолеть разрыв между молекулярной структурой и физиологическим поведением, необходимо понимание динамики атомного разрешения калиевых каналов.
Из-за высокого сходства последовательностей между порами KcsA и другими белками эукариотических K-ионных каналов, KcsA предоставил важную информацию о поведении других важных белков, проводящих напряжение, таких как дрозофиллы, производные Shaker, и калиевый канал человека hERG. KcsA использовался в исследованиях мутагенеза для моделирования взаимодействий между hERG и различными лекарственными соединениями. Такие тесты могут выявить взаимодействия между лекарственным средством и каналом hERG, которые вызывают синдром приобретенного удлиненного интервала QT, необходимы для определения сердечной безопасности новых лекарств. Кроме того, модели гомологии, основанные на кристаллической структуре KcsA в замкнутом состоянии, были сгенерированы с помощью вычислений для построения представления множественных состояний сердечного K-канала hERG. Такие модели показывают гибкость канала hERG и могут последовательно предсказывать сродство связывания набора различных лигандов, взаимодействующих с ионным каналом. Анализ сложных структур лиганд-hERG может быть использован для управления синтезом аналогов лекарств с пониженной активностью hERG на основе структуры лекарственного средства и потенциала стыковки.