
Войти
эндосомальные сортировочные комплексы, необходимые для транспортировки (ESCRT ) аппарат состоит из цитозольных белковых комплексов, известных как ESCRT-0, ESCRT-I, ESCRT-II и ESCRT-III. Вместе с рядом дополнительных белков эти комплексы ESCRT обеспечивают уникальный режим ремоделирования мембраны, в результате чего мембраны изгибаются / отталкиваются от цитоплазмы. Эти компоненты ESCRT были выделены и изучены у ряда организмов, включая дрожжи и людей.
Механизм ESCRT играет жизненно важную роль в ряде клеточных процессов, включая мультивезикулярное тело (MVB) биогенез, клеточная абсциссия и вирусное почкование. Биогенез мультивезикулярных тел (MVB) - это процесс, в котором белки, меченные убиквитином, проникают в органеллы, называемые эндосомами, посредством образования везикул. Этот процесс необходим клеткам для разрушения неправильно свернутых и поврежденных белков. Без механизма ESCRT эти белки могут накапливаться и вызывать нейродегенеративное заболевание. Например, аномалии компонентов ESCRT-III могут приводить к неврологическим расстройствам, таким как (HSP). Абсциссия клеток, процесс, посредством которого отщепляется мембрана, соединяющая две дочерние клетки, также опосредуется механизмом ESCRT. Без комплексов ESCRT дочерние клетки не могут разделиться, и будут образовываться аномальные клетки, содержащие вдвое больше ДНК. Эти клетки неизбежно будут разрушены в результате процесса, известного как апоптоз. Наконец, почкование вируса или процесс, посредством которого определенные типы вирусов выходят из клеток, могут не происходить в отсутствие аппарата ESCRT. Это неизбежно предотвратит распространение вирусов от клетки к клетке.
 Краткое описание механизмов и вспомогательных белков ESCRT.
Краткое описание механизмов и вспомогательных белков ESCRT. Каждый из комплексов ESCRT и дополнительных белков имеет уникальные структуры, которые обеспечивают различные биохимические функции. Для каждого белкового компонента аппарата ESCRT существует ряд синонимов как для дрожжей, так и для многоклеточных животных. Сводная таблица всех этих белков представлена ниже.
У дрожжей существуют следующие комплексы / дополнительные белки:
Комплекс ESCRT-0 играет жизненно важную роль в образовании мультивезикулярных телец посредством связывание и кластеризацию убиквитинированных белков и / или рецепторов на поверхности клетки. Затем комплекс отвечает за связывание с липидом на эндосомальной мембране, который привлекает эти меченые белки к эндосоме. После правильной локализации эти белки затем попадают в эндосому через везикулы, образуя мультивезикулярные тельца, и в конечном итоге доставляются в лизосому, где они разрушаются. Этот процесс важен, поскольку он является основным путем деградации поврежденных белков, прошедших через Гольджи. Компоненты комплекса ESCRT-0 существуют следующим образом:
Комплекс представляет собой 1: 1 гетеродимер Vps27 (вакуолярный протеин, сортирующий протеин 27) и Hse1. Vps27 и Hse1 димеризуют от до антипараллельные спиральные домены GAT (названные так в честь белков GGA и Tom1). И Vps27, и Hse1 содержат амино-терминал домен VHS (названный так, потому что он содержится в V ps27, H RS и S ТАМ белки). Эти домены VHS связывают убиквитин с белками, которые клетка стремится разрушить. Убиквитин также может ассоциироваться с мотивами, взаимодействующими с убиквитином, такими как мотив Hse1 или двусторонний домен, обнаруженный на Vps27. Домен FYVE (названный в честь четырех белков, в которых он был первоначально идентифицирован: Fab1p, YOTB, Vac1 и EEA1) находится между доменами взаимодействующих мотивов VHS и убиквитина Vps27. Фосфатидилинозитол 3 -фосфат, обычный эндосомальный липид, связывается с этим доменом FYVE, что приводит к привлечению ESCRT-0 к эндосоме.
Роль ESCRT-I комплекс должен способствовать образованию мультивезикулярных телец за счет кластеризации убиквитинированных белков и выступая в качестве моста между комплексами ESCRT-0 и ESCRT-II. Он также играет роль в распознавании и ремоделировании мембраны во время отсоединения мембраны, образуя кольца по обе стороны от среднего тела делящихся клеток. ESCRT-I также отвечает за рекрутирование ESCRT-III, который формирует зону сжатия непосредственно перед разделением клеток. Более того, ESCRT-I играет роль в почковании вируса, взаимодействуя со специфическими вирусными белками, что приводит к привлечению дополнительных механизмов ESCRT к потенциальному месту высвобождения вируса. Детали механизма ESCRT-I описаны ниже.
Комплекс ESCRT-I представляет собой гетеротетрамер (1: 1: 1: 1) Vps23, Vps28, Vps37 и Mvb12. Собранный гетеротетрамер выглядит как стержнеобразный стержень, состоящий из Vps23, Vps37 и Mvb12 с веерообразным колпачком, состоящим из одиночных спиралей из Vps23, Vps28 и Vps37. Vps23 содержит один вариантный домен убиквитина E2, который отвечает за связывание убиквитина, комплекса ESCRT-0, и с мотивом PTAP (pролин, tхреонин, aланин, pролин) вирусного Gag белки. Сразу после этого варианта домена убиквитина E2 присутствует мотив, богатый пролином (GPPX 3 Y), который направляет ESCRT-I к среднему телу во время абсциссии мембраны. Mvb12 также может связывать убиквитин через свой карбокси-конец. Vps28 отвечает за взаимодействие ESCRT-I и ESCRT-II, связываясь с доменом GLUE (G RAM- L ик U связывание биквитина в E AP45) Vps36 через его карбокси-концевой домен пучка с четырьмя спиралями.
Комплекс ESCRT-II функционирует в основном во время биогенез мультивезикулярных телец и доставка белков, меченных убиквитином, в эндосому. Белки, меченные убиквитином, передаются от ESCRT-0 к ESCRT-I, а затем к ESCRT-II. ESCRT-II связывается с ESCRT-III, который зажимает груз, содержащий везикулу, закрытым. Специфические аспекты ESCRT-II следующие:
ESCRT-II представляет собой гетеротетрамер (2: 1: 1), состоящий из двух субъединиц Vps25, одной Vps22 и одной Субблок Vps36. Молекулы Vps25 содержат мотивы PPXY, которые связываются с мотивами крылатой спирали (WH) Vps22 и Vps36, создавая Y-образный комплекс с Vps22 и Vps36 в качестве основания и молекулами Vps25 в качестве плеч. Молекулы Vps25 также содержат мотивы WH, которые отвечают за взаимодействие ESCRT-II с ESCRT-III. Vps36 содержит домен GLUE, который связывает фосфатидилинозитол-3-фосфат и Vps28 ESCRT-I. Два домена цинковых пальцев вплетены в домен GLUE дрожжевого Vps36. Один из этих доменов с цинковыми пальцами связывает карбокси-концевой домен Vps28, а другой связывается с убиквитином.
Комплекс ESCRT-III, вероятно, является наиболее важным из всех ESCRT. механизм, потому что он играет роль во всех процессах, опосредованных ESCRT. Во время отрыва мембраны и образования почки вируса ESCRT-III формирует длинные филаменты, которые наматываются вокруг места сужения мембраны непосредственно перед разрывом мембраны. Это опосредование опорожнения происходит посредством взаимодействий с комплексом centralspindlin. Эти нитевидные структуры также присутствуют во время формирования мультивезикулярных тел и функционируют как кольцеобразный забор, который закупоривает почкующиеся пузырьки, чтобы предотвратить утечку грузовых белков в цитозоль клетки. ESCRT-III существует и функционирует следующим образом:
Комплекс ESCRT-III отличается от всех других механизмов ESCRT тем, что он существует только временно и содержит как существенные, так и второстепенные компоненты. Основные субъединицы должны быть собраны в правильном порядке (Vps20, Snf7, Vps24, затем Vps2) для функционирования оборудования. Несущественные субъединицы включают Vps60, Did2 и Ist1. Vps20 инициирует сборку ESCRT-III, действуя как зародышеобразователь сборки полимера Snf7. Затем Vps24 связывается с Snf7, чтобы ограничить комплекс и задействовать Vps2. Затем Vps2 добавляет в комплекс Vps4. Все «свободные» цитозольные формы каждой субъединицы считаются закрытыми. То есть карбокси-концевая часть каждой субъединицы складывается сама по себе автоингибирующим образом, стабилизируя мономерные субъединицы. Карбокси-конец большинства субъединиц ESCRT-III, как основных, так и несущественных, содержит MIM (M IT (микротрубочка взаимодействующий и транспортный домен) i взаимодействующий м отиф) мотивы. Эти мотивы отвечают за связывание Vps4 и AAA-ATPase спастин.
Белки Vps4-Vta1 необходимы для удаления других компонентов ESCRT (обычно ESCRT-III) с мембран после завершения определенного процесса. Существуют некоторые дискуссии относительно того, отщепляет ли Vps4 комплекс ESCRT-III или ремоделирует комплекс, так что один компонент отщепляется в определенное время. Считается, что Vta1 действует как активатор Vps4, помогая его сборке и повышая его активность AAA-ATPase. Эти белки функционируют следующим образом:
субъединицы Vps4 имеют два функциональных домена, аминоконцевой домен MIT и центральный домен AAA-ATPase. Домен MIT отвечает за взаимодействие Vps4 с доменом MIM Vps2. Домен AAA-ATPase гидролизует ATP, чтобы привести в действие разборку комплекса ESCRT-III. Такое «удаление» ESCRT-III позволяет повторно использовать все связанные субъединицы для дальнейшего использования. Vta1 - это димерный белок, содержащий один домен VSL (названный так, потому что он содержится в белках V ps4, S BP1 и L IP5), который обеспечивает связывание с Vps4, и домен MIT для связывания с субъединицей ESCRT-III Vps60. Хотя это и не обязательно, Vta1 помогает в сборке кольца Vps4, ускоряет АТФазную активность Vsp4 и способствует разборке ESCRT-III.
Основная функция Bro1 - рекрутировать деубиквитиназ в комплекс ESCRT-III. Это приводит к удалению убиквитиновых меток из белков, нацеленных на деградацию в лизосомах, непосредственно перед образованием мультивезикулярных телец. Также предполагалось, что Bro1 помогает стабилизировать ESCRT-III, в то время как убиквитиновые метки отщепляются от грузовых белков.
Bro1 содержит аминоконцевой домен Bro1, который связывается с Snf7 ESCRT-III. Это связывание приводит Bro1 к месту отрыва мембраны. Bro1 также связывает каталитический домен Doa4, убиквитингидролазу (деубиквитиназу), перемещая его к месту отрыва. Doa4 удаляет убиквитин из грузовых белков, нацеленных на лизосомы.
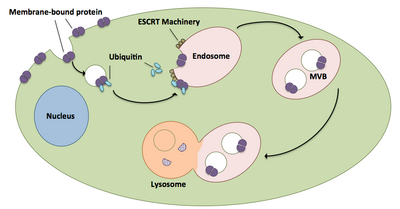 Перенос мембраносвязанных белков в лизосому с использованием аппарата ESCRT. Связанные с мембраной белки попадают в клетку через эндоцитоз. Теги убиквитина на белке распознаются аппаратом ESCRT и рекрутируются в эндосому. Образуются мультивезикулярные тельца, которые затем сливаются с лизосомой, где эти белки расщепляются. Адаптировано из.
Перенос мембраносвязанных белков в лизосому с использованием аппарата ESCRT. Связанные с мембраной белки попадают в клетку через эндоцитоз. Теги убиквитина на белке распознаются аппаратом ESCRT и рекрутируются в эндосому. Образуются мультивезикулярные тельца, которые затем сливаются с лизосомой, где эти белки расщепляются. Адаптировано из. Множественные тельца играют большую роль в транспорте убиквитинированных белков и рецепторов к лизосомам. Комплексы ESCRT транспортируют убиквитинированный груз к клеточным пузырькам, которые отращиваются непосредственно в эндосомный компартмент клетки, образуя мультивезикулярные тела. Эти мультивезикулярные тела в конечном итоге сливаются с лизосомами, вызывая деградацию груза. Более подробное описание процесса, включая связанный с ним механизм, существует следующим образом:
 Рекрутирование комплексов ESCRT в среднее тело. Cep-55 связывает MKLP1. Cep-55 нанимает ESCRT-I и ALIX. ESCRT-I и ALIX набирают ESCRT-III. ESCRT-III образует спираль вокруг шейки мембраны между дочерними клетками, что приводит к сжатию и расщеплению. Адаптировано из.
Рекрутирование комплексов ESCRT в среднее тело. Cep-55 связывает MKLP1. Cep-55 нанимает ESCRT-I и ALIX. ESCRT-I и ALIX набирают ESCRT-III. ESCRT-III образует спираль вокруг шейки мембраны между дочерними клетками, что приводит к сжатию и расщеплению. Адаптировано из. Отслоение мембраны во время цитокинеза - это процесс, посредством которого мембрана, соединяющая две дочерние клетки, расщепляется во время деления клетки. Поскольку он консервативен у ряда архей, расслоение мембраны считается самой ранней ролью механизма ESCRT. Процесс начинается, когда центросомный белок CEP55 рекрутируется в среднее тело делящихся клеток в ассоциации с MKLP1, митотическим кинезином -подобным белком, который ассоциируется с микротрубочками. Затем CEP55 задействует субъединицу Vps23 ESCRT-I и дополнительный белок ALIX, которые образуют кольца по обе стороны от среднего тела. ESCRT-I и ALIX рекрутируют ESCRT-III через его субъединицу Snf7. Субъединицы ESCRT-III Vps20, Snf7, Vps24, Vps2 и Did2 образуют спиралевидные фибриллы, прилегающие к кольцам, образованным Vps23. Образование этой спиралевидной структуры деформирует мембрану, и спастин ААА-АТФазы вводится с помощью Did2 и Ist1, чтобы расщепить микротрубочки, образованные в среднем теле. Vps4 затем катализирует разборку комплекса ESCRT-III, в результате чего образуются две вновь разделенные дочерние клетки. Процесс расслоения мембраны был описан с использованием белков многоклеточных животных, поскольку этот процесс в большей степени изучен у многоклеточных животных.
 Ретровирусное отпочкование ВИЧ. A) Накопление вирусных белков под клеточной мембраной заставляет вирус выступать наружу. Б) Сужение образуется комплексами ESCRT в основании выступа мембраны, вызывая образование везикулы, содержащей вирус. C) Почка отщипывается, оставляя свободный внеклеточный вирион. (Фотография предоставлена доктором Мэтью Гонда (Wikimedia Commons: ноябрь 1998 г.), ID изображения Национального института рака: 2382)
Ретровирусное отпочкование ВИЧ. A) Накопление вирусных белков под клеточной мембраной заставляет вирус выступать наружу. Б) Сужение образуется комплексами ESCRT в основании выступа мембраны, вызывая образование везикулы, содержащей вирус. C) Почка отщипывается, оставляя свободный внеклеточный вирион. (Фотография предоставлена доктором Мэтью Гонда (Wikimedia Commons: ноябрь 1998 г.), ID изображения Национального института рака: 2382) Высвобождение вирусных частиц, также известное как почкование вируса, - это процесс свободные вирионы высвобождаются из клеток посредством захвата механизма ESCRT клетки-хозяина. Ретровирусы, такие как ВИЧ-1 и человеческие Т-лимфотропные вирус, а также ряд вирусов в оболочке, включая вирус Эбола, требуют оборудования ESCRT для выхода из клетки-хозяина. Этот процесс инициируется вирусными белками Gag, основными структурными белками ретровирусной оболочки, которые взаимодействуют с TSG101 комплекса ESCRT-I и дополнительным белком ALIX. Субъединицы ESCRT-III (существенными являются только CHMP4 и CHMP2) рекрутируются в место зарождения вируса, чтобы сузить и перерезать шейку зачатка аналогично тому, как описано для обрыва мембраны во время цитокинеза. Затем Vps4 рециркулирует компоненты ESCRT-III в цитозоль, и вирус высвобождается из клетки. Механизм, описанный здесь, использует белки многоклеточных животных, так как вирусное почкование более широко изучено у многоклеточных животных.