
Войти
 Чарльз Дарвин в 1868 году
Чарльз Дарвин в 1868 году Научное исследование видообразования - как виды эволюционируют, чтобы стать новыми видами - началось примерно во времена Чарльза Дарвина в середине XIX века. Многие натуралисты в то время признали взаимосвязь между биогеографией (способом распространения видов) и эволюцией видов. В 20-м веке область видообразования расширилась, и основные участники, такие как Эрнст Майр, исследовали и задокументировали географические закономерности и взаимосвязи видов. Эта область приобрела известность с современным эволюционным синтезом в начале того века. С тех пор исследования видообразования значительно расширились.
Язык видообразования стал более сложным. Споры по поводу схем классификации механизмов видообразования и репродуктивной изоляции продолжаются. В 21 веке наблюдается возрождение исследований видообразования с появлением новых методов, таких как молекулярная филогенетика и систематика. Видообразование в основном подразделяется на дискретные режимы, которые соответствуют скорости потока генов между двумя зарождающимися популяциями. Однако сегодня исследования привели к разработке альтернативных схем и открытию новых процессов видообразования.
 Единственная фигура в книге Дарвина 1859 года О происхождении видов, древо родословных, разделившееся на новые виды
Единственная фигура в книге Дарвина 1859 года О происхождении видов, древо родословных, разделившееся на новые виды Чарльз Дарвин представил идею о том, что виды могут развиваться и разделяться на отдельные линии, ссылаясь на это как на спецификацию в своей книге 1859 года О происхождении Виды. Только в 1906 году современный термин видообразование был введен биологом Оратором Ф. Куком. Дарвин в своей публикации 1859 года сосредоточился в первую очередь на изменениях, которые могут произойти внутри вида, и меньше на том, как виды могут делиться на две части. Почти повсеместно признано, что книга Дарвина напрямую не касалась своего названия. Вместо этого Дарвин считал, что видообразование происходит в результате проникновения видов в новые экологические ниши.
Существуют разногласия относительно того, Чарльз Дарвин признал истинную географическую модель видообразования в своей публикация «Происхождение видов». В главе 11 «Географическое распространение» Дарвин обсуждает географические барьеры для миграции, заявляя, например, что «барьеры любого рода или препятствия на пути к свободной миграции тесно и важным образом связаны с различиями между продукцией различных регионов [ мира] ". Ф. Дж. Саллоуэй утверждает, что позиция Дарвина по видообразованию была, по меньшей мере, «вводящей в заблуждение» и, возможно, позже дезинформировала Вагнера и Дэвида Старра Джордана, заставив их поверить в то, что Дарвин рассматривал симпатрическое видообразование как наиболее важный способ видообразования. Тем не менее, Дарвин так и не принял полностью концепцию Вагнера о географическом видообразовании.
биолог-эволюционист Джеймс Маллет утверждает, что мантра, повторяемая в книге Дарвина «Происхождение видов», в которой никогда не обсуждались видообразование, является надуманной. Заявление началось с Томаса Генри Хаксли и Джорджа Романеса (современники Дарвина), которые заявили, что Дарвин не смог объяснить происхождение непостоянства и бесплодия гибридов. Подобные утверждения были обнародованы мутационистской школой мысли в конце 20-го века и даже после современного эволюционного синтеза Ричарда Гольдшмидта. Еще один сильный сторонник этого взгляда на Дарвина пришел из Майра. Майр утверждал, что Дарвин не смог решить проблему видообразования, поскольку он не определял виды, используя концепцию биологических видов. Однако мнение Майра не было полностью принято, поскольку записные книжки Дарвина по трансмутации содержали записи о роли изоляции в разделении видов. Более того, многие идеи Дарвина о видообразовании в значительной степени совпадают с современными теориями адаптивной радиации и экологического видообразования.
 Немецкий путешественник Мориц Вагнер (1813 г.) –1887)
Немецкий путешественник Мориц Вагнер (1813 г.) –1887) Признание географических факторов, влияющих на популяции видов, присутствовало еще до Дарвина, и многие натуралисты осознавали роль изоляции во взаимоотношениях видов. В 1833 году К. Л. Глогер опубликовал «Изменчивость птиц под влиянием климата», в которой он описал географические вариации, но не признал, что географическая изоляция была индикатором прошлых событий видообразования. Другой естествоиспытатель в 1856 г., Волластон, изучал островных жуков в сравнении с видами на материке. Он видел в изоляции ключ к их различию. Однако он не осознавал, что эта модель возникла из-за видообразования. Один натуралист Леопольд фон Бух (1825) распознал географические закономерности и прямо заявил, что географическая изоляция может привести к разделению видов на новые. Майр предполагает, что фон Бух, вероятно, был первым натуралистом, который действительно предположил географическое видообразование. Другие натуралисты, такие как Генри Уолтер Бейтс (1863), признали и приняли закономерности как свидетельство видообразования, но в случае Бэйта не предложили последовательной модели.
В 1868 году Мориц Вагнер был первым, кто предложил концепцию географического видообразования, в которой он использовал термин Теория разделения. Эдвард Бэгнал Поултон, биолог-эволюционист и убежденный сторонник важности естественного отбора, подчеркнул роль географической изоляции в продвижении видообразования, в процессе создания термина «симпатрическое видообразование» в 1904 году.
Вагнер и другие естествоиспытатели, изучавшие географическое распределение животных, такие как Карл Джордан и Дэвид Старр Джордан, заметил, что близкородственные виды часто были географически изолированы друг от друга (аллопатрически распределены), что привело к отстаиванию важности географической изоляции в происхождении видов. Считается, что Карл Джордан признал объединение мутации и изоляции в происхождении новых видов, что резко контрастировало с преобладающими в то время взглядами. Дэвид Старр Джордан повторил предложение Вагнера в 1905 году, предоставив множество свидетельств природы в поддержку теории и заявив, что географическая изоляция очевидна, но, к сожалению, была проигнорирована большинством генетиков и экспериментальных эволюционных биологов в то время. Джоэл Асаф Аллен предложил называть наблюдаемую схему географического разделения близкородственных видов «законом Иордании» (или законом Вагнера). Несмотря на утверждения, большинство систематиков приняли географическую модель видообразования.
Многие из первых терминов, используемых для описания видообразования, были выделены Эрнстом Майром. Он был первым, кто изложил современную тогда литературу в своей публикации 1942 года Систематика и происхождение видов с точки зрения зоолога и в своей последующей публикации 1963 года. Как и работы Джордана, они основывались на прямых наблюдениях за природой, документируя возникновение географического видообразования. Он описал три режима: географический, полугеографический и негеографический; которые сегодня называются аллопатрическими, парапатрическими и симпатрическими соответственно. Публикация Майра 1942 года, на которую сильно повлияли идеи Карла Джордана и Поултона, считалась авторитетным обзором видообразования на протяжении более 20 лет - и до сих пор ценна по сей день.
Основное внимание в работах Майра уделялось важности географии в облегчении видообразования; острова часто выступают центральной темой многих выдвигаемых концепций видообразования. Одним из них была концепция перипатрического видообразования, варианта аллопатрического видообразования (с тех пор он различил эти два модуса, называя их перипатрическим и дихопатрическим). Эта концепция возникла в результате интерпретации теории разделения Вагнера как формы эффекта основателя видообразования, сосредоточенного на небольших географически изолированных видах. Позднее эта модель была расширена и модифицирована, чтобы включить половой отбор в 1976 и 1980 годах.
Многие генетики в то время мало что сделали, чтобы преодолеть разрыв между генетика естественного отбора и происхождение репродуктивных барьеров между видами. Рональд Фишер предложил модель видообразования в своей публикации 1930 года Генетическая теория естественного отбора, где он описал действие разрушающего отбора на симпатрических или парапатрических популяциях - с репродуктивной изоляцией, дополненной подкреплением. Другие генетики, такие как Дж. Б.С. Холдейн даже не признал, что виды были реальными, в то время как Сьюэлл Райт проигнорировал эту тему, несмотря на принятие аллопатрического видообразования.
Основные участники включения видообразования в современный эволюционный синтез были Эрнст Майр и Феодосий Добжанский. Добжанский, генетик, в 1937 году опубликовал Генетика и происхождение видов, в которой сформулировал генетическую основу того, как может происходить видообразование. Он признал, что видообразование было нерешенной проблемой в биологии в то время, отвергая позицию Дарвина о том, что новые виды возникли в результате занятия новых ниш - утверждая, что репродуктивная изоляция вместо этого основана на барьерах для потока генов. Впоследствии Майр провел обширную работу по географии видов, подчеркнув важность географического разделения и изоляции, в которой он заполнил пробелы Добжанского относительно происхождения биоразнообразия (в его книге 1942 года). Обе их работы породили, не без противоречий, современное понимание видообразования; стимулирование обширных исследований по этой теме. Кроме того, это распространялось на растения и животных с G. Книга Ледьярда Стеббинса Вариация и эволюция растений и гораздо более поздняя книга 1981 года, написанная Верном Грантом.
 Эрнстом Майром, влиятельным эволюционным биологом
Эрнстом Майром, влиятельным эволюционным биологом В 1947 году на конференции Принстонского университета «был достигнут консенсус между генетиками, палеонтологами и систематиками, и что эволюционная биология была учреждена как независимая биологическая дисциплина». Этот синтез 20 века включал видообразование. С тех пор эти идеи постоянно и неоднократно подтверждались.
После синтеза исследования видообразования продолжались в основном в рамках естествознания и биогеографии - с гораздо меньшим упором на генетику. Изучение видообразования пережило самый большой рост с 1980-х годов с притоком публикаций и множеством новых терминов, методов, концепций и теорий. Этот «третий этап» работы - как Джерри А. Койн и Х. Аллен Орр сказал, что это привело к усложнению языка, используемого для описания многих процессов видообразования. Исследования и литература по видообразованию стали «огромными, разрозненными и все более техническими».
С 1980-х годов новые исследовательские инструменты повысили надежность исследования, чему способствовали новые методы, теоретические основы, модели и подходы.. Койн и Орр обсуждают современные разработки после 1980-х годов, сосредоточенные вокруг пяти основных тем:
Экологи осознали, что экологические факторы, лежащие в основе видообразования были недостаточно представлены. Это привело к росту исследований, касающихся роли экологии в облегчении видообразования - справедливо обозначенного экологического видообразования. Это внимание к экологии привело к появлению множества новых терминов, связанных с препятствиями для воспроизводства (например, аллохроническое видообразование, при котором поток генов уменьшается или удаляется по времени периодов размножения; или изоляция среды обитания, в которой виды занимают разные среды обитания в пределах одного и того же района). Симпатрическое видообразование, которое Майр считает маловероятным, стало широко признанным. Исследования влияния нату Разросся рациональный отбор на видообразование, включая процесс подкрепления.
Исследователи давно обсуждают роль полового отбора, естественного отбора и генетического дрейф в видообразовании. Дарвин подробно обсуждал половой отбор, а его работы значительно расширил Рональд Фишер; однако только в 1983 г. биолог Мэри Джейн Уэст-Эберхард осознала важность полового отбора в видообразовании. Естественный отбор играет роль в том, что любой отбор, направленный на репродуктивную изоляцию, может привести к видообразованию - прямо или косвенно. Генетический дрейф широко изучается с 1950-х годов, особенно с помощью моделей видообразования с пиковым сдвигом в результате генетического дрейфа. Майр отстаивал эффекты основателя, в которых изолированные особи, подобные тем, что встречаются на островах недалеко от материка, испытывают сильное узкое место в популяции, поскольку они содержат лишь небольшую выборку генетической изменчивости в основной популяции. Позже другие биологи, такие как Хэмптон Л. Карсон, Алан Темплтон, Сергей Гаврилец и Алан Гастингс, разработали родственные модели видообразования генетический дрейф, отмечая, что острова были населены в основном эндемичными видами. Роль отбора в видообразовании широко поддерживается, в то время как видообразование с эффектом основателя - нет, поскольку подвергалось ряду критических замечаний.
 Видообразование представлено как континуум потока генов, где
Видообразование представлено как континуум потока генов, где  равняется скорости обмена генами. В этом континууме могут существовать три основных географических способа видообразования (аллопатрический, парапатрический и симпатрический ), а также другие негеографические формы.
равняется скорости обмена генами. В этом континууме могут существовать три основных географических способа видообразования (аллопатрический, парапатрический и симпатрический ), а также другие негеографические формы. На протяжении всей истории исследований, касающихся видообразования, классификации и разграничения видов и процессов обсуждались. Джулиан Хаксли разделил видообразование на три отдельных вида: географическое видообразование, генетическое видообразование и экологическое видообразование. Сьюэлл Райт предложил десять различных режимов. Эрнст Майр отстаивал важность физического, географического разделения популяций видов, считая его важным для видообразования. Первоначально он предложил три основных режима, известных сегодня: географический, полугеографический, негеографический; соответствующие аллопатрической, парапатрической и симпатрической соответственно.
Фраза «способы видообразования» определена неточно, чаще всего это указывает на видообразование, происходящее в результате географического распространения вида. Более кратко, современная классификация видообразования часто описывается как происходящая в континууме потока генов (т. Е. Аллопатрия на 

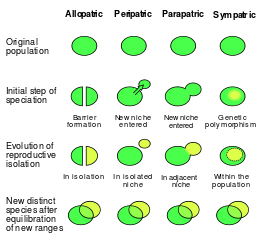 Сравнение трех классических географических способов видообразования: аллопатрический, парапатрический и симпатрический ; перипатрическое видообразование включено как частный случай аллопатрического видообразования.
Сравнение трех классических географических способов видообразования: аллопатрический, парапатрический и симпатрический ; перипатрическое видообразование включено как частный случай аллопатрического видообразования. По мере того, как исследования расширялись на протяжении десятилетий, географическая схема подвергалась сомнению. Одни исследователи считают традиционную классификацию устаревшей, а другие отстаивают ее достоинства. Сторонники негеографических схем часто оправдывают негеографические классификации не отрицанием важности репродуктивной изоляции (или даже самих процессов), а тем фактом, что она упрощает сложность видообразования. Одна из основных критических замечаний по поводу географической структуры состоит в том, что она произвольно разделяет биологический континуум на прерывистые группы. Другая критика связана с тем фактом, что, когда видообразование рассматривается как континуум потока генов, парапатрическое видообразование оказывается необоснованно представленным целым континуумом - с аллопатричным и симпатрическим существованием в крайностях. Койн и Орр утверждают, что схема географической классификации ценна тем, что биогеография контролирует силу действующих эволюционных сил, поскольку поток генов и география четко связаны. Джеймс Маллет и его коллеги утверждают, что дихотомия между симпатией и аллопатрией важна для определения степени, в которой естественный отбор влияет на видообразование. Киркпатрик и Равинье классифицируют видообразование с точки зрения его генетической основы или сил, управляющих репродуктивной изоляцией. Здесь географические способы видообразования классифицируются как типы ассортивного спаривания. Фитцпатрик и его коллеги считают, что биогеографическая схема «является отвлекающим фактором, который может ввести в заблуждение, если настоящая цель - понять влияние естественного отбора на дивергенцию». Они утверждают, что для полного понимания видообразования необходимо изучить «пространственные, экологические и генетические факторы», участвующие в дивергенции. Сара Виа признает важность географии в видообразовании, но предлагает отказаться от классификации по этой схеме.
Симпатрическое видообразование с самого начала с Дарвином (который не придумал этот термин) было спорным вопросом. Майр, наряду со многими другими биологами-эволюционистами, истолковал взгляд Дарвина на видообразование и происхождение биоразнообразия как возникновение видов, вступающих в новые экологические ниши - форму симпатрического видообразования. До Майра симпатрическое видообразование считалось основным способом видообразования. В 1963 году Майр выступил с резкой критикой, сославшись на различные недостатки теории. После этого симпатрическое видообразование перестало быть популярным среди биологов, и лишь недавно к ним вернулся интерес. Некоторые биологи, такие как Джеймс Маллет, полагают, что взгляд Дарвина на видообразование был неправильно истолкован и истолкован Майром. Сегодня симпатрическое видообразование подтверждается данными лабораторных экспериментов и наблюдений на природе.
На протяжении большей части истории видообразования гибридизация (полиплоидия) была спорным вопросом, как утверждают ботаники. и зоологи традиционно по-разному рассматривали роль гибридизации в видообразовании. Карл Линней был первым, кто предположил гибридизацию в 1760 году, Ойвинд Виндж был первым, кто подтвердил аллополиплоидию в 1917 году, а позже был проведен эксперимент. Клаузен и Гудспид в 1925 году подтвердили выводы. Сегодня это широко признано как распространенный механизм видообразования.
Исторически зоологи считали гибридизацию редким явлением, а ботаники считали ее обычным явлением для видов растений. Ботаники Г. Ледьярд Стеббинс и Верн Грант были двумя известными ботаниками, которые отстаивали идею гибридного видообразования в 1950-1980-х годах. Гибридное видообразование, также называемое полиплоидным видообразованием (или полиплоидией), - это видообразование, которое возникает в результате увеличения количества наборов хромосом. По сути, это форма симпатрического видообразования, которое происходит мгновенно. Грант ввел термин в 1981 г.; особая форма гибридного видообразования, при которой новый вид является результатом гибридизации и репродуктивно изолирован от обоих родителей. В последнее время биологи все чаще признают, что гибридное видообразование может происходить и у животных.
 Молодой естествоиспытатель Альфред Рассел Уоллес в 1862 году
Молодой естествоиспытатель Альфред Рассел Уоллес в 1862 году Концепция видообразования методом подкрепления имеет сложную историю, и его популярность среди ученых со временем значительно изменилась. Теория подкрепления пережила три фазы исторического развития:
Первоначально он был предложен Альфредом Расселом Уоллесом в 1889 году и получил название «эффект Уоллеса» - термин, который сегодня редко используется учеными. Гипотеза Уоллеса отличалась от современной концепции тем, что фокусировалась на постзиготической изоляции, усиленной групповым отбором. Добжанский был первым, кто дал полное современное описание процесса в 1937 году, хотя сам термин не был введен до 1955 года У. Фрэнк Блэр.
В 1930 году Рональд Фишер представил первое генетическое описание процесса подкрепления в Генетической теории естественного отбора, а в 1965 и 1970 годах - первое компьютерное моделирование. были запущены, чтобы проверить его достоверность. Позже были проведены популяционно-генетические и количественные генетические исследования, показавшие, что совершенно непригодные гибриды приводят к увеличению презиготической изоляции. После того, как идея Добжанского вышла на передний план исследований видообразования, она получила значительную поддержку - Добжанский предположил, что она иллюстрирует последний этап видообразования (например, после того, как аллопатрическая популяция входит во вторичный контакт ). В 1980-х годах многие биологи-эволюционисты начали сомневаться в правдоподобности этой идеи, основываясь не на эмпирических данных, а в основном на росте теории, которая считала это маловероятным механизмом репродуктивной изоляции. Тогда возник ряд теоретических возражений. С начала 1990-х годов популярность подкрепления возродилась, и биологи-эволюционисты признали ее правдоподобность - главным образом благодаря внезапному увеличению объема данных, эмпирическим данным лабораторных исследований и исследований природы, сложному компьютерному моделированию и теоретической работе.
Научный язык, касающийся подкрепления, также менялся с течением времени: разные исследователи применяли разные определения этого термина. Впервые использованное для описания наблюдаемых различий в брачных криках у лягушек Gastrophryne внутри гибридной зоны вторичного контакта, подкрепление также использовалось для описания географически разделенных популяций, которые испытывают вторичный контакт. отделяет неполную постзиготическую изоляцию от полной изоляции, ссылаясь на неполную изоляцию как на подкрепление, а на полностью изолированные популяции как на смещение репродуктивного характера. считалось, что смещение репродуктивного признака представляет либо ассортивное спаривание, либо расхождение признаков для распознавания партнера (особенно между симпатрическими популяциями). Согласно этому определению, это включает презиготическое расхождение и полную постзиготическую изоляцию. и Мохамед Нур рассматривают любое обнаруженное увеличение предзиготической изоляции как подкрепление, если это является реакцией на отбор против спаривания между двумя разными видами. Койн и Орр утверждают, что «истинное усиление ограничено случаями, в которых усилена изоляция между таксонами, которые все еще могут обмениваться генами».