
Войти
| Lassa mammarenavirus | |
|---|---|
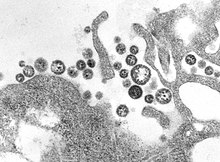 | |
| TEM микрофотография Lassa mammarenavirus вирионы | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Класс: | Ellioviricetes |
| Порядок: | Bunyavirales |
| Семейство: | Arenaviridae |
| Род: | Mammarenavirus |
| Вид: | Lassa mammarenavirus |
| Синонимы | |
| |
Lassa mammarenavirus (LASV ) - это аренавирус, вызывающий геморрагическую лихорадку Ласса, разновидность вирусной геморрагической лихорадки (VHF), у людей и других приматов. Lassa mammarenavirus - это развивающийся вирус и агент выбора, требующий сдерживания, эквивалентного 4-му уровню биобезопасности. Он эндемичен в странах Западной Африки, особенно в Сьерра-Леоне, Гвинейской Республике, Нигерии и Либерии, где ежегодная заболеваемость зараженности составляет от 300 000 до 500 000 случаев, что приводит к 5 000 смертей в год.
По состоянию на 2012 год открытия в регионе реки Мано на западе Африки расширили эндемичный зона между двумя известными эндемичными регионами Ласса, что указывает на то, что LASV более широко распространен в тропической лесистой саванне экозоне в Западной Африке. Нет утвержденных вакцин против лихорадки Ласса для использования у людей.
В 1969 году миссионерская медсестра Лаура Вайн заболела загадочной болезнью, которой заразился акушерский пациент в Ласса, деревне в Борно. Штат, Нигерия. Затем она была доставлена в Джос, Нигерия, где и умерла. Впоследствии заразились еще двое, одна из которых была пятидесятидвухлетней медсестрой Лили Пиннео, которая ухаживала за Лорой Вайн. Образцы из Pinneo были отправлены в Йельский университет в Нью-Хейвене, где Jordi Casals <впервые выделил новый вирус, который позже будет известен как Lassa mammarenavirus, Соня Бакли и другие. Казальс заболел лихорадкой и чуть не умер; один техник умер от этого. К 1972 году было обнаружено, что крыса с несколькими половыми членами, Mastomys natalensis, была основным резервуаром вируса в Западной Африке, способной выделять вирус с мочой и фекалиями, не проявляя видимых симптомов.
 Структура и геном вируса Ласса
Структура и геном вируса Ласса Вирусы Ласса представляют собой оболочечные, одноцепочечные, двусегментированные вирусы самбисенсой РНК. Их геном состоит из двух сегментов РНК, каждый из которых кодирует по два белка, по одному в каждом смысле, всего четыре вирусных белка. Большой сегмент кодирует небольшой белок цинковый палец (Z), который регулирует транскрипцию и репликацию, и РНК-полимеразу (L). Небольшой сегмент кодирует нуклеопротеин (NP) и поверхностный предшественник гликопротеина (GP, также известный как вирусный спайк), который протеолитически расщепляется на гликопротеины оболочки GP1 и GP2, которые связываются к рецептору альфа-дистрогликана и опосредуют проникновение в клетку-хозяин.
Лихорадка Ласса вызывает геморрагическую лихорадку, часто проявляемую иммуносупрессией. Lassa mammarenavirus очень быстро реплицируется и демонстрирует временный контроль репликации. Первым этапом репликации является транскрипция мРНК копий отрицательного или отрицательного смысла генома. Это обеспечивает адекватное снабжение вирусными белками для последующих стадий репликации, поскольку белки NP и L транслируются с мРНК. Затем геном с положительным или положительным смыслом создает копии вирусной комплементарной РНК. Копии РНК являются матрицей для получения потомства с негативным смыслом, но из него также синтезируется мРНК. МРНК, синтезированная из vcRNA, транслируется с образованием белков GP и Z. Такой временный контроль позволяет продуцировать спайковые белки в последнюю очередь и, следовательно, задерживать распознавание иммунной системой хозяина.
Нуклеотидные исследования генома показали, что Ласса имеет четыре линии происхождения: три найдены в Нигерии, а четвертая - в Гвинее, Либерии и Сьерра-Леоне. Похоже, что нигерийские штаммы были предками других, но для подтверждения этого требуется дополнительная работа.
 Механизмы проникновения аренавирусов Старого и Нового Света.
Механизмы проникновения аренавирусов Старого и Нового Света. Вирус Lassa mammarenavirus получает доступ к клетка-хозяин с помощью рецептора клеточной поверхности альфа-дистрогликан (альфа-DG), универсального рецептора белков внеклеточного матрикса. Он разделяет этот рецептор с прототипом аренавируса Старого Света вируса лимфоцитарного хориоменингита. Распознавание рецептора зависит от конкретной сахарной модификации альфа-дистрогликана группой гликозилтрансфераз, известных как БОЛЬШИЕ белки. Конкретные варианты генов, кодирующих эти белки, по-видимому, проходят положительный отбор в Западной Африке, где Ласса является эндемиком. Альфа-дистрогликан также используется в качестве рецептора вирусами аренавирусов клады С Нового Света (вирусы Oliveros и Latino). Напротив, аренавирусы Нового Света кладов A и B, которые включают важные вирусы Machupo, Junin и Sabia в дополнение к непатогенному вирусу Amapari. используйте рецептор трансферрина 1. Небольшая алифатическая аминокислота в позиции 260 аминокислоты гликопротеина GP1 требуется для высокоаффинного связывания с альфа-DG. Кроме того, положение 259 аминокислоты GP1 также представляется важным, поскольку все аренавирусы, демонстрирующие высокоаффинное связывание альфа-DG, содержат объемную ароматическую аминокислоту (тирозин или фенилаланин) в этом положении.
В отличие от большинства вирусов в оболочке. которые используют покрытые клатрином ямки для проникновения в клетки и связываются со своими рецепторами в зависимости от pH, Ласса и вирус лимфоцитарного хориоменингита вместо этого используют эндоцитотический путь, независимый от клатрина, кавеолин, динамин и актин. Попав внутрь клетки, вирусы быстро доставляются к эндосомам посредством везикулярного транспорта, хотя и в значительной степени независимого от небольших GTPases и. При контакте с эндосомой происходит pH-зависимое слияние мембран, опосредованное гликопротеином оболочки, который при более низком pH эндосомы связывает лизосомный белок LAMP1, что приводит к слиянию мембран и выходу из эндосомы.
 Жизненный цикл вируса Ласса
Жизненный цикл вируса Ласса Жизненный цикл маммаренавируса Ласса аналогичен аренавирусам Старого Света. Маммаренавирус Lassa проникает в клетку посредством рецептор-опосредованного эндоцитоза. Какой эндоцитотический путь используется, пока неизвестно, но, по крайней мере, вход в клетки чувствителен к истощению холестерина. Сообщалось, что интернализация вируса ограничивается при истощении холестерина. Рецептор, используемый для входа в клетку, представляет собой альфа- дистрогликан, высококонсервативный и повсеместно экспрессируемый рецептор клеточной поверхности для белков внеклеточного матрикса. Дистрогликан, который позже расщепляется на альфа-дистрогликан, и бета-дистрогликан первоначально экспрессируется в большинстве клеток в зрелые ткани и обеспечивает молекулярную связь между ЕСМ и цитоскелетом на основе актина. После того, как вирус проникает в клетку посредством эндоцитоза, опосредованного альфа-дистрогликаном, среда с низким pH запускает pH-зависимое слияние мембран и высвобождает комплекс RNP (вирусный рибонуклеопротеин) в цитоплазму. Вирусная РНК распаковывается, а репликация и транскрипция инициируются в цитоплазме. Когда начинается репликация, геномы S и L РНК синтезируют антигеномные S и L РНК, а из антигеномных РНК синтезируются геномные S и L РНК. И геномная, и антигеномная РНК необходимы для транскрипции и трансляции. S-РНК кодирует белки GP и NP (вирусный нуклеокапсидный белок), а L-РНК кодирует белки Z и L. Белок L, скорее всего, представляет собой вирусную РНК-зависимую РНК-полимеразу. Когда клетка инфицирована вирусом, L-полимераза связывается с вирусной РНП и инициирует транскрипцию геномной РНК. 5 ’и 3’ концевые 19-нуклеотидные участки вирусного промотора обоих сегментов РНК необходимы для распознавания и связывания вирусной полимеразы. Первичная транскрипция сначала транскрибирует мРНК из геномных S- и L-РНК, которые кодируют белки NP и L соответственно. Транскрипция заканчивается в структуре «стебель-петля» (SL) внутри межгеномной области. Аренавирусы используют стратегию отрывания кэпа для получения кэп-структур из клеточных мРНК, и это опосредуется эндонуклеазной активностью L-полимеразы и связывающей кэп активностью NP. Антигеномная РНК транскрибирует вирусные гены GPC и Z, кодируемые в геномной ориентации, из сегментов S и L. соответственно. Антигеномная РНК также служит матрицей для репликации. После трансляции GPC он посттрансляционно модифицируется в эндоплазматическом ретикулуме. GPC расщепляется на GP1 и GP2 на более поздней стадии секреторного пути. Сообщалось, что за это расщепление отвечает клеточная протеаза SKI-1 / S1P. Расщепленные гликопротеины включаются в оболочку вириона, когда вирус прорастает и высвобождается из клеточной мембраны.
лихорадка Ласса чаще всего вызывается маммаренавирусом Ласса. Симптомы включают гриппоподобное заболевание, характеризующееся лихорадкой, общей слабостью, кашлем, болью в горле, головной болью и желудочно-кишечными проявлениями. Геморрагические проявления включают проницаемость сосудов.
Попадая, маммаренавирус Ласса поражает почти все ткани человеческого тела. Он начинается с слизистой оболочки, кишечника, легких и мочевыделительной системы, а затем переходит в сосудистую систему.
Основными мишенями вируса являются антигенпрезентирующие клетки, в основном дендритные клетки ) и эндотелиальные клетки. В 2012 году сообщалось, что нуклеопротеин (NP) вируса маммаренавируса Ласса саботирует реакцию врожденной иммунной системы хозяина. Обычно, когда патоген проникает в хозяина, система врожденной защиты распознает патоген-ассоциированные молекулярные паттерны (PAMP) и активирует иммунный ответ. Один из механизмов обнаруживает двухцепочечную РНК (дцРНК), которая синтезируется только вирусами с отрицательным смыслом. В цитоплазме рецепторы дцРНК, такие как RIG-I (ген I, индуцируемый ретиноевой кислотой) и MDA-5 (ген 5, связанный с дифференцировкой меланомы), обнаруживают дцРНК и инициируют сигнальные пути, которые перемещают IRF-3 (интерферон регуляторный фактор 3) и другие факторы транскрипции в ядро. Транслоцированные факторы транскрипции активируют экспрессию интерферонов 𝛂 и, и они инициируют адаптивный иммунитет. NP, кодируемый в Lassa mammarenavirus, играет важную роль в вирусной репликации и транскрипции, но он также подавляет врожденный ответ IFN хозяина, ингибируя транслокацию IRF-3. Сообщается, что NP маммаренавируса Ласса обладают активностью экзонуклеазы только по отношению к дцРНК. активность экзонуклеазы дцРНК NP противодействует ответам IFN путем переваривания PAMP, что позволяет вирусу уклоняться от иммунных ответов хозяина.