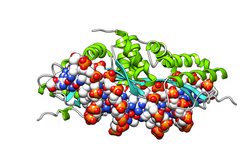
Кристаллическая структура
I-CreI связана с его
последовательностью распознавания ДНК. В
фермент связывается в виде гомодимера; одна субъединица изображена желтым, другая - розовым. Фермент показан на поверхности; Молекула ДНК представлена в виде набора сфер, каждая из которых окрашена в соответствии с ее
химическим элементом.
В ГСНЕ эндонуклеазы представляют собой набор эндонуклеаз, кодируемых либо в виде отдельно стоящие гены внутри интронов, так как слитые белков с принимающими, или в качестве самостоятельного сплайсинга интеины. Они катализируют гидролиз геномной ДНК в клетках, которые их синтезируют, но делают это в очень немногих или даже в отдельных местах. Ремонт гидролизованной ДНК клеткой-хозяином часто приводит к тому, что ген, кодирующий хоминг-эндонуклеазу, копируется в сайт расщепления, отсюда термин «хоминг» для описания движения этих генов. Таким образом, самонаводящиеся эндонуклеазы могут передавать свои гены горизонтально в популяции хозяина, увеличивая частоту своих аллелей с большей, чем менделевской, скоростью.
СОДЕРЖАНИЕ
- 1 Происхождение и механизм
- 2 Номенклатура
- 3 Сравнение с рестрикционными ферментами
- 4 Структурные семейства
- 5 Доменная архитектура
- 6 См. Также
- 7 ссылки
- 8 Внешние ссылки
Происхождение и механизм
Хотя происхождение и функция самонаводящихся эндонуклеаз все еще исследуются, наиболее устоявшаяся гипотеза рассматривает их как эгоистичные генетические элементы, подобные транспозонам, поскольку они способствуют сохранению генетических элементов, которые их кодируют, независимо от предоставления функционального атрибута организму-хозяину..
Последовательности распознавания самонаводящейся эндонуклеазы достаточно длинные, чтобы происходить случайным образом только с очень низкой вероятностью (примерно один раз в 7 × 10 9 п.н. ) и обычно встречаются в одном или очень нескольких экземплярах на геном. Как правило, благодаря механизму самонаведения ген, кодирующий эндонуклеазу (HEG, «ген самонаводящейся эндонуклеазы»), находится в последовательности узнавания, которую расщепляет фермент, тем самым прерывая последовательность узнавания самонаводящейся эндонуклеазы и ограничивая разрезание ДНК только теми сайтами, которые не (пока) несут HEG.
Перед передачей один аллель несет ген (HEG +), а другой - нет (HEG -), и поэтому он подвержен разрезанию ферментом. Как только фермент синтезируется, он разрушает хромосому в HEG - аллеле, инициируя ответ системы репарации клеточной ДНК. Повреждение восстанавливается с помощью рекомбинации, по образцу противоположного неповрежденного аллеля ДНК, HEG +, который содержит ген эндонуклеазы. Таким образом, ген копируется на аллель, у которого его изначально не было, и передается из поколения в поколение. Этот процесс называется «самонаведение».
Номенклатура
Самонаводящиеся эндонуклеазы всегда обозначаются префиксом, который идентифицирует их геномное происхождение, за которым следует дефис: «I-» для самонаводящихся эндонуклеаз, кодируемых внутри интрона, «PI-» (для «белковой вставки») для тех, которые кодируются внутри интеина. Некоторые авторы предложили использовать префикс «F-» («автономный») для вирусных ферментов и других природных ферментов, не кодируемых интронами или интеинами, и «H-» («гибрид») для ферментов, синтезированных в лаборатории. Затем трехбуквенное имя получается из биноминального названия организма, взяв одну заглавную букву из названия рода и две строчные буквы из конкретного названия. (Некоторое смешивание обычно выполняется для гибридных ферментов.) Наконец, римская цифра обозначает разные ферменты, обнаруженные в одном и том же организме:
- PI-TliII ( P30317 ) - это второй идентифицированный фермент, кодируемый интеином, обнаруженным в архее Thermococcus litoralis.
- H-DreI ( PDB : 1MOW ) - первая синтетическая эндонуклеаза самонаведения, созданная в лаборатории из ферментов I-DmoI ( P21505 ) и I-CreI ( P05725 ), взятых соответственно из Desulfurococcus mobilis и Chlamydomonas reinhardtii.
Сравнение с рестрикционными ферментами
Самонаводящиеся эндонуклеазы отличаются от ферментов рестрикции типа II по нескольким параметрам:
- В то время как ферменты рестрикции типа II связываются с короткими, обычно симметричными, узнавающими последовательностями размером от 4 до 8 п.н., хомящие эндонуклеазы связываются с очень длинными и во многих случаях асимметричными последовательностями распознавания, охватывающими от 12 до 40 п.н.
- Самонаводящиеся эндонуклеазы обычно более толерантны к заменам в последовательности узнавания. Незначительные изменения в последовательности узнавания обычно снижают активность хоминг-эндонуклеаз, но часто не отменяют ее полностью, как это часто происходит с рестрикционными ферментами.
- У самонаводящихся эндонуклеаз общие структурные мотивы, которые предполагают, что существует четыре семейства, в то время как невозможно определить просто узнаваемые и различимые семейства рестрикционных ферментов типа II.
- Самонаводящиеся эндонуклеазы действуют как мономеры или гомодимеры и часто требуют ассоциированных белков для регулирования их активности или образования рибонуклеопротеидных комплексов, где РНК является неотъемлемым компонентом каталитического аппарата. Ферменты рестрикции типа II могут также функционировать отдельно, как мономеры или гомодимеры, или с дополнительными субъединицами белка, но дополнительные субъединицы отличаются от субъединиц самонаводящихся эндонуклеаз. Таким образом, для их действия могут потребоваться субъединицы ограничения, модификации и специфичности.
- Наконец, самонаводящиеся эндонуклеазы имеют более широкое филогенетическое распределение, встречающееся во всех трех биологических доменах - архей, бактерий и эукарий. Ферменты рестрикции типа II встречаются только у архей, бактерий и некоторых вирусов. Самонаводящиеся эндонуклеазы также экспрессируются во всех трех компартментах эукариотической клетки: ядрах, митохондриях и хлоропластах. Открытые рамки считывания, кодирующие самонаводящиеся эндонуклеазы, были обнаружены в интронах, интеинах и в автономной форме между генами, тогда как гены, кодирующие гены рестрикционных ферментов типа II, были обнаружены только в автономной форме, почти всегда в тесной связи с генами, кодирующими родственные ферменты, модифицирующие ДНК. Таким образом, хотя рестрикционные ферменты типа II и хоминг-эндонуклеазы разделяют функцию расщепления двухцепочечной ДНК, они, по-видимому, эволюционировали независимо.
Структурные семьи
В настоящее время известно шесть структурных семейств. Их консервативные структурные мотивы :
- LAGLIDADG: Каждый полипептид имеет 1 или 2 мотива LAGLIDADG. Последовательность LAGLIDADG представляет собой консервативную последовательность аминокислот, где каждая буква представляет собой код, идентифицирующий конкретный остаток. Эта последовательность напрямую участвует в процессе разрезания ДНК. Те ферменты, у которых есть только один мотив, работают как гомодимеры, создавая седло, которое взаимодействует с большой бороздкой каждого полусайта ДНК. Мотивы LAGLIDADG вносят аминокислотные остатки как в интерфейс белок-белок между доменами или субъединицами белков, так и в активные центры фермента. Ферменты, которые обладают двумя мотивами в одной белковой цепи, действуют как мономеры, создавая седло аналогичным образом. Первые структуры, которые должны были быть определены для самонаводящихся эндонуклеаз (PI-SceI и I-CreI, о которых было сообщено в 1997 г.), были из структурного семейства LAGLIDADG. к его сайту-мишени ДНК также сообщалось.
- GIY-YIG: они имеют только один мотив GIY-YIG в N-концевой области, который взаимодействует с ДНК в участке разреза. Фермент-прототип этого семейства - I-TevI, который действует как мономер. Сообщалось об отдельных структурных исследованиях ДНК-связывающих и каталитических доменов I-TevI, первый из которых связан с его ДНК-мишенью, а второй - в отсутствие ДНК.
- His-Cys box ( Pfam PF05551 ): эти ферменты содержат область из 30 аминокислот, которая включает 5 консервативных остатков: два гистидина и три цистеина. Они координируют катион металла, необходимый для катализа. I-PpoI является наиболее охарактеризованным ферментом этого семейства и действует как гомодимер. О его структуре было сообщено в 1998 году. Возможно, он связан с семейством HNH, поскольку у них есть общие черты.
- HNH: ( Pfam CL0263 ): они имеют консенсусную последовательность примерно из 30 аминокислот. Он включает две пары консервативных гистидинов и одну аспарагин, которые создают домен цинкового пальца. I-HmuI ( P34081 ) - это наиболее охарактеризованный фермент этого семейства, который действует как мономер. Его структура была опубликована в 2004 году ( PDB : 1U3E ).
- PD- (D / E) xK ( Pfam CL0236 ): Эти ферменты содержат каталитический домен канонической нуклеазы, обычно обнаруживаемый в эндонуклеазах рестрикции типа II. Наиболее охарактеризованный фермент этого семейства, I-Ssp6803I ( Q57253 ), действует как тетрамер. Его структура была опубликована в 2007 году ( PDB : 2OST ). Общая складка консервативна во многих семействах эндонуклеаз, все из которых принадлежат к суперсемейству PD- (D / E) xK.
- Vsr-подобный / EDxHD (DUF559, InterPro : IPR007569 ): эти ферменты были обнаружены в Глобальной базе данных метагеномных проб океана и впервые описаны в 2009 году. Термин «Vsr-подобный» относится к наличию С-концевого домена нуклеазы, который отображает узнаваемая гомология бактериальных эндонуклеаз репарации очень коротких участков (Vsr). Структура была решена в 2011 году, что подтвердило гомологию Vsr. Считается частью суперсемейства PD- (D / E) xk.
Доменная архитектура
Хоминг-эндонуклеаза дрожжей PI-Sce представляет собой эндонуклеазу типа LAGLIDADG, кодируемую как интеин, который сплайсируется из другого белка ( P17255 ). Структура высокого разрешения выявляет два домена : эндонуклеолитический центр, напоминающий С-концевой домен белков Hedgehog, и домен Hint (Hedgehog / Intein), содержащий активный сайт сплайсинга белка.
Смотрите также
использованная литература
внешние ссылки
- Perler FB. «InBase». Архивировано из оригинала на 2010-08-02. Проверено 9 августа 2010.
База данных и реестр Intein (из биолабораторий Новой Англии)
- Perler FB (январь 2002 г.). «InBase: база данных Intein». Nucleic Acids Res. 30 (1): 383–4. DOI : 10.1093 / NAR / 30.1.383. PMC 99080. PMID 11752343.
Эта статья включает текст из общественного достояния
Pfam и
InterPro :
IPR007868 Эта статья включает текст из общественного достояния
Pfam и
InterPro :
IPR007869 
 кристаллическая структура минипредшественника пи-скеи
кристаллическая структура минипредшественника пи-скеи