
Войти
В В биохимии добавление формильной функциональной группы называется формилированием . Формильная функциональная группа состоит из карбонила, связанного с водородом. При присоединении к R-группе, Формильная группа называется ed и альдегид.
 Формильная функциональная группа показана синим цветом.
Формильная функциональная группа показана синим цветом. Формилирование было идентифицировано в нескольких важных биологических процессах. Метионин был впервые обнаружен в кишечной палочке Маркером и Сэнгером в 1964 году как формилированный, а позже было установлено, что он участвует в инициации синтеза белка в бактериях и органеллах. Образование N-формилметионина катализируется ферментом метионил-тРНК трансформилазой. Кроме того, в биосинтезе пуринов de novo происходят две реакции формилирования. Эти реакции катализируются ферментами глицинамидрибонуклеотид (GAR) трансформилаза и 5-аминоимидазол-4-карбоксиамидриботид (AICAR) трансформилаза. Совсем недавно было обнаружено, что формилирование представляет собой модификацию гистона, которая может модулировать экспрессию гена.
Формилирование относится к любым химическим процессам, в которых соединение функционализируется формильной группой (-CH = O). В органической химии этот термин чаще всего используется в отношении ароматических соединений (например, преобразование бензола в бензальдегид в Гаттерманне – Кохе реакция ). В биохимии реакция катализируется ферментами, такими как формилтрансферазы.
Формилирование метанофурана инициирует метаногенез цикл. Формильная группа образована из диоксида углерода и преобразуется в метан.
 Цикл для метаногенеза, демонстрируя начальное формилирование метанофурана.
Цикл для метаногенеза, демонстрируя начальное формилирование метанофурана. В бактериях и органеллах о начале синтеза белка сигнализирует образование формилметионил-тРНК (тРНК). Эта реакция зависит от 10-формилтетрагидрофолата и фермента метионил-тРНК формилтрансферазы. Эта реакция не используется эукариотами или археями, так как присутствие тРНК в небактериальных клетках считается интрузивным материалом и быстро устраняется. После образования тРНК доставляется к 30S-субъединице рибосомы, чтобы начать синтез белка. fMet имеет ту же последовательность кодонов, что и метионин. Однако fMet используется только для инициации синтеза белка и, таким образом, обнаруживается только на N-конце белка. Во время остального перевода используется метионин. В E.coli тРНК специфически распознается фактором инициации IF-2, поскольку формильная группа блокирует образование пептидной связи на N-конце метионина.
После завершения синтеза белка формильная группа метионина может быть удалена с помощью пептид-деформилазы. Остаток метионина может быть дополнительно удален ферментом метионинаминопептидазой.
 Химический синтез N-формилметионина катализируется ферментом метионил-тРНК формилтрансферазой
Химический синтез N-формилметионина катализируется ферментом метионил-тРНК формилтрансферазой Два формилирования реакции необходимы на одиннадцати стадиях синтеза de novo инозинмонофосфата (IMP), предшественника пуриновых рибонуклеотидов AMP и GMP. Глицинамидрибонуклеотид (GAR) трансформилаза катализирует формилирование GAR до формилглицинамидинриботида (FGAR) в четвертой реакции пути. На предпоследнем этапе биосинтеза пурина de novo риботид 5-аминоимидазол-4-карбоксиамида (AICAR) формилируется в 5-формаминоимидазол-4-карбоксамид риботид (FAICAR) с помощью трансформилазы AICAR.
PurN GAR трансформилаза обнаружена у эукариот и прокариот. Однако вторая трансформилаза GAR, PurT GAR трансформилаза, была идентифицирована в E. coli. Хотя два фермента не имеют консервативной последовательности и требуют разных доноров формила, удельная активность и Km для GAR одинаковы как для PurT, так и для PurN-GAR трансформилазы.
PurN GAR трансформилаза 1CDE использует кофермент N10-формилтетрагидрофолат (N10-формил-THF) в качестве донора формила для формилирования α-аминогруппы GAR. У эукариот трансформилаза PurN GAR является частью большого многофункционального белка, но обнаруживается как отдельный белок у прокариот.
Предполагается, что реакция формилирования происходит посредством реакции прямого переноса, в которой аминогруппа GAR нуклеофильно атакует N10-формил-THF с образованием тетраэдрического промежуточного соединения. Поскольку α-аминогруппа GAR относительно реакционноспособна, предполагается, что депротонирование нуклеофила происходит под действием растворителя. В активном сайте Asn 106, His 108 и Asp 144 позиционируются, чтобы способствовать переносу формила. Однако исследования мутагенеза показали, что эти остатки по отдельности не важны для катализа, поскольку только мутации двух или более остатков ингибируют фермент. Основываясь на структуре, предполагается, что отрицательно заряженный Asp144 увеличивает pKa His108, позволяя протонированной имидазолиевой группе His108 повышать электрофильность формильной группы N10-формил-THF. Кроме того, считается, что His108 и Asn106 стабилизируют оксианион, образующийся в переходном состоянии.
PurT GAR трансформилазы требует формиата в качестве донора формила и АТФ для катализа. Было подсчитано, что трансформилаза PurT GAR осуществляет 14-50% формилирования GAR в E. coli. Фермент является членом суперсемейства белков, захватывающих АТФ.
Для трансформилазы PurT GAR был предложен последовательный механизм, в котором предполагается, что короткоживущий промежуточный формилфосфат сначала образует. Этот промежуточный формилфосфат затем подвергается нуклеофильной атаке амином GAR для переноса формильной группы. Промежуточный формилфосфат был обнаружен в экспериментах по мутагенезу, в которых мутантная трансфоримилаза PurT GAR имела слабое сродство к формиату. Инкубация трансформилазы PurT GAR с формилфосфатом, ADP и GAR дает как ATP, так и FGAR. Это дополнительно указывает на то, что формилфосфат может быть промежуточным продуктом, поскольку он кинетически и химически компетентен для проведения реакции формилирования в ферменте. Ферментный фосфатный промежуточный продукт, предшествующий формилфосфатному промежуточному продукту, также был предложен для образования на основе исследований позиционного изотопного обмена. Однако структурные данные показывают, что формиат может быть расположен для прямой атаки на γ-фосфат АТФ в активном центре фермента с образованием промежуточного формилфосфата.
Трансформилазе AICAR требуется кофермент N10-формилтетрагидрофолат (N10-формил-ТГФ) в качестве донора формила для формилирования AICAR в FAICAR. Однако трансформилаза AICAR и трансформилаза GAR не обладают высоким сходством последовательностей или структурной гомологией.
Амин на AICAR намного менее нуклеофильный, чем его аналог на GAR, из-за делокализации электронов в AICAR посредством конъюгации. Следовательно, для протекания реакции формилирования необходимо активировать нуклеофил N5 в AIRCAR. Было обнаружено, что гистидин 268 и лизин 267 необходимы для катализа и консервативны во всех трансформилазе AICAR. Гистидин 268 участвует в депротонировании нуклеофила N5 AICAR, в то время как лизин 267, как предполагается, стабилизирует тетраэдрический промежуточный продукт.
 Формилирование - это происходящая посттрансляционная модификация на остатки лизина.
Формилирование - это происходящая посттрансляционная модификация на остатки лизина. ε-Формилирование является одной из многих посттрансляционных модификаций, которые происходят на гистоновых белках, которые, как было показано, модулируют конформации хроматина и активацию генов.
 Формилирование лизина может конкурировать с ацетилированием как посттрансляционная модификация.
Формилирование лизина может конкурировать с ацетилированием как посттрансляционная модификация. Формилирование было идентифицировано на Nε остатков лизина в гистонах и белках. Эта модификация наблюдалась в линкерных гистонах и белках группы с высокой подвижностью, она очень распространена и, как полагают, играет роль в эпигенетике функции хроматина. Было показано, что формилированные лизины играют роль в связывании ДНК. Кроме того, формилирование было обнаружено на лизинах гистонов, которые, как известно, также ацетилированы и метилированы. Таким образом, формилирование может блокировать другие посттрансляционные модификации. Формилирование наиболее часто выявляется на 19 различных сайтах модификации гистона H1. Генетическая экспрессия клетки сильно нарушена формилированием, что может вызвать такие заболевания, как рак. Развитие этих модификаций может быть связано с окислительным стрессом.
В гистоновых белках лизин обычно модифицируется гистоновыми ацетилтрансферазами (HAT) и гистоновыми деацетилазами (HDAC или KDAC). Ацетилирование лизина имеет фундаментальное значение для регуляции и экспрессии определенных генов. Окислительный стресс создает совершенно иную среду, в которой ацетил-лизин может быть быстро вытеснен образованием формил-лизина из-за высокой реакционной способности форм формилфосфата. В настоящее время считается, что эта ситуация вызвана окислительным повреждением ДНК. Был предложен механизм образования формилфосфата, который в значительной степени зависит от окислительно поврежденной ДНК и в основном управляется радикальной химией внутри клетки. Полученный формилфосфат затем можно использовать для формилирования лизина. Считается, что оксидативный стресс играет роль в доступности остатков лизина на поверхности белков и в возможности их формилирования.
 Формилфосфат является предполагаемым продуктом окислительного повреждения ДНК.
Формилфосфат является предполагаемым продуктом окислительного повреждения ДНК. Ингибирование ферментов, участвующих в биосинтезе пуринов, использовалось как средство потенциальная лекарственная мишень для химиотерапии.
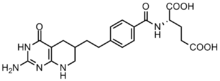 Химическая структура лометрексола.
Химическая структура лометрексола. Раковые клетки требуют высоких концентраций пуринов для облегчения деления и имеют тенденцию полагаться на синтез de novo, а не на путь восстановления нуклеотидов. Несколько ингибиторов на основе фолиевой кислоты были разработаны для ингибирования реакций формилирования под действием трансформилазы GAR и трансформилазы AICAR. Первый ингибитор трансформилазы GAR, лометрексол [(6R) 5,10-дидеазатетрагидрофолат], был разработан в 1980-х годах в результате сотрудничества Eli Lilly и академических лабораторий.
Хотя по структуре он похож на N10- формил-ТГФ, лометрексол не способен проводить реакции переноса одного углерода. Кроме того, были синтезированы несколько ингибиторов трансформилазы на основе GAR. Было обнаружено, что разработка ингибиторов на основе фолиевой кислоты является особенно сложной задачей, поскольку ингибиторы также подавляют фермент фолиполиглутаматсинтазу, который добавляет дополнительные γ-глутаматы к моноглутаматным фолатам и антифолатам после попадания в клетку для повышения сродства к ферменту. Это повышенное сродство может привести к устойчивости к антифолатам.
Синдром Ли - это нейродегенеративное заболевание, которое было связано с дефектом в реакции ферментативного формилирования. Синдром Ли обычно связан с дефектами окислительного фосфорилирования, которое происходит в митохондриях. Секвенирование экзома было использовано для выявления мутации в гене, кодирующем митохондриальную метионил-тРНК-формилтрансферазу (MTFMT) у пациентов с лихорадкой Ли. синдром. Считается, что мутация c.626C>T, выявленная в MTFMT, приводящая к симптомам синдрома Ли, изменяет сплайсинг экзонов, приводя к мутации сдвига рамки считывания и преждевременному стоп-кодону. Было обнаружено, что индивидуумы с мутацией MTFMT c.626C>T имеют сниженные уровни fMet-tRNAMet и изменения в уровне формилирования митохондрически транслируемого COX1. Эта ссылка предоставляет доказательства необходимости формилированного метионина в инициации экспрессии определенных митохондриальных генов.