
Войти
| Amphidromus | |
|---|---|
 | |
| Вид Amphidromus roseolabiatus имеет правую спиральную оболочку. | |
 | |
| Вид Amphidromus fuscolabris имеет левостороннюю спиральную оболочку. | |
| Научная классификация | |
| Домен: | Eukaryota |
| Царство: | Animalia |
| Тип: | Mollusca |
| Класс: | Gastropoda |
| Подкласс: | Heterobranchia |
| Надотряд: | Eupulmonata |
| Порядок: | Stylommatophora |
| Инфраотряд: | |
| Надсемейство: | Helicoidea |
| Семейство: | Camaenidae |
| Род: | Amphidromus. Альберс, 1850 |
| Типовой вид | |
| Amphidromus perversus. (Линней,) | |
| Вид | |
См. Текст | |
| Разнообразие | |
| более 110 видов | |
Амфидромус - это род тропических дышащих воздухом наземных улиток, наземных лёгких брюхоногих моллюсков моллюски из семейства Camaenidae. Раковины Amphidromus относительно большие, от 25 мм (0,98 дюйма) до 75 мм (3,0 дюйма) в максимальном размере, и особенно красочные. В XVIII веке они были одними из первых индонезийских раковин наземных улиток, Европу путешественниками и исследователями. С тех пор этот период род активно изучается: в с начала 19 до середины 20 веков естествоиспытатели и зоологи написали несколько всеобъемлющих монографий и каталогов. Современные исследования на лучшее понимание эволюционных отношений внутри группы, а также решение таксономических проблем.
Род Amphidromus необычен тем, что он включает виды, которые имеют правое свертывание раковины, и которые имеют левое свертывание раковины. Кроме того, некоторые виды этого рода особенно примечательны тем, что в их популяции одновременно входят особи двусторонней и правосторонней намотки раковины. Это крайне редкое явление, очень интересное биологам. Исследования, посвященные мягкой анатомии Amphidromus, разрознены и фрагментарны. Информация о внутренней анатомии известна только по нескольким видам, и более крупных сравнительных морфологических исследований никогда не проводилось.
Виды рода Amphidromus древесные - другими словами, это древесные улитки. Однако более подробной информации об их привычках пока нет. Общие пищевые привычки этих улиток неизвестны, но некоторые виды питаются микроскопическими грибами, лишайниками или наземными водорослями. На самих амфидромов охотятся птицы, змеи и, вероятно, также более мелкие млекопитающие, такие как крысы.
 Немецкий зоолог Эдуард фон Мартенс (1831– 1904 г. опубликовал первую всеобъемлющую монографию по роду Amphidromus в 1867 году, многие из его концепций все еще используются.
Немецкий зоолог Эдуард фон Мартенс (1831– 1904 г. опубликовал первую всеобъемлющую монографию по роду Amphidromus в 1867 году, многие из его концепций все еще используются. родовое название происходит от древнегреческих слов amphí (ἀμφί), что означает «с обеих сторон», и drómos (δρόμος), что означает «бег», называясь на различные хиральности оболочек. Раковины Amphidromus относительно большие и довольно красочные; значительное количество из них были первых индонезийских раковин наземных улиток, привезенных в Европу путешественниками и исследователями в 18. Для сравнения, малакологи собрали гораздо меньшее количество экземпляров.
Некоторые виды и формы были мощи до 1800 г., большинство из них не имеют адекватных данных о местонахождении. По крайней мере, два названия - Amphidromus laevus (Müller, 1774) и форма А. perversus ф. aureus Martyn, 1784 - до сих пор (по состоянию на 2017 год) еще не зарегистрированы с точного местонахождения. В первой половине девятнадцатого века были названы многие виды разновидностей, опять же, как правило, с плохими данными о местонахождении. Только после того, как Эдуард фон Мартенс (1867) опубликовал свою монографию, не было предпринято попытка охватить весь комплекс этого рода. Монография 1867 г. содержала значительный объем информации как о вариациях внутри рода, так и о проблемах географического распространения вида. Многие концепции, возникшие у фон Мартенса, до сих пор (по состоянию на 2017 год) используются.
В 1896 году 142 конкретных названия и названия сортов были в восемь групп видов, объединены в общей сложности 64 вида. Когда появилась монография Генри Августа Пилсбри 1900 года Руководство по конхологии, количество видов в роде Amphidromus увеличилось до 81, и они были разделены на девятнадцать групп. Исследование Пилсбри осталось единственной иллюстрированной монографией этого рода, и оно до сих пор считается незаменимым для любого серьезного изучения рода.
С 1900 года основные таксономические исследования по Amphidromus были фаунистический (изучение фауны определенной территории или района) по объему. Работы американского малаколога Пола Барча (1917, 1918, 1919) о филиппинских лучше, Бернхарда Ренша (1932) о формах Малых Зондских островов и (1950, 1959) о популяциях Явана и Суматры. Возможно, наиболее ценный вклад принадлежит (1921), который обсуждал вариацию в пределах A. Contrarius и на Тимор ; вариации цвета и формы были хорошо проиллюстрированы на серии цветных пластин.
Литература, опубликованная после 1900 года, создает множество разрозненных описаний новых цветовых форм и подвидов. Из 309 имен в номенклатурном списке 111 (35,9%) были опубликованы после Пилсбри (1900). (1953) перечислил типовые образцы в Зенкенбергском музее и проиллюстрировали многие ранее не изображенные виды. Фрэнк Фортескью Лейдлоу Алан Солем (1961) распознал 74 вида по названию и посчитал, что материал с островов Банда, вероятно, представляет собой неописанный вид. Одиннадцать из видов, признанных Laidlaw Solem, были возможности после выхода монографии Пилсбри. Однако несколько видов, признанных Пилсбри, могут быть подчинены подвидовому или сортовому статусу, а некоторые названия были перенесены в incertae sedis, поскольку они основаны на ссылках столетней давности, которые не были подтверждены более поздними коллекционерами. Фактически, исследование Laidlaw Solem (1961) является дополнением к монографии Пилсбри с его обширными пластинами, и многие выводы Laidlaw Solem относительно взаимоотношений цветовых форм, описываемых видов, взяты не столько из новых образцов, сколько из степени изменчивости, которая была обозначена Ханиэлем (1921) в его пионерском исследовании.
Виды рода Amphidromus обычно имеют гладкую, глянцевую, ярко окрашенную, удлиненную или коническую форму, правую или правую сторону. левосторонне свернутые гильзы. Раковины умеренно большие, от 25 мм (0,98 дюйма) до 70 мм (2,8 дюйма) в максимальном размере, имеют от 6 до 8 выпуклых оборотов. Их цветовой узор обычно однотонный желтоватый или зеленоватый, но может быть разнообразным. Отверстие имеет наклонную илицевидную форму, без зубцов или складок, высота отверстия составляет от двух пятых до одной трети общей высоты оболочки. Перистом расширен и / или отражен, иногда утолщен. колумелла может быть прямой или изогнутой, а теменная мозоль от слабой до хорошо развитой, а пупок может быть открытым или закрытым. радула лопатчатая, с зубцами, расположенными рядами, обычно с монокоспидными центральными зубом и двустворчатыми или трехстворчатыми боковыми зубцами. челюсть тонкая и слабая, с низкими плоскими ребрами. Паллиальная область сигмуретрозная с очень длинной узкой почкой. гениталии - это гениталии типичных каменид, с длинным семеприемником, коротким пенисом с низким прикреплением ретракционной мышцы и короткой или длинной эпифаллической слепой кишкой (жгутик и аппендикс). сперматофоры имеют пятиугольный контур в поперечном сечении. Amphidromus - это обычно древесные животные.
раковины Amphidromus относительно большие, от одного до трех дюймов в высоту и красочные. Амфидром имеет продолговато-коническую или яйцевидно-коническую спиралевидную оболочку с 5-8 оборотами. Раковина может быть тонкой и хрупкой или очень тяжелой и прочной, без известной корреляции структуры раковины с распространением или средой среды обитания.
У некоторых видов этого рода витки оболочки неизменно вправо, а во многих других точно так же неизменно слева. Однако значительным числом видов этого рода являются «амфидром »; Этот термин означает, что и левосторонняя, и правосторонняя намотка оболочки проявляется в одной и той же популяции. Можно сказать, что они являются «полиморфными » для направления наматывания оболочки, но существует только два типа наматывания оболочки, они запуск как «диморфные » при намотке. Два типа скручивания раковины встречаются у некоторых видов в равном количестве, у других видов отчетливо преобладает одна фаза. Пока нет информации о наследственности этого признака у Амфидром.
Потому что почти все другие виды амфидромовыххоногих моллюсков, например, принадлежащие к родам Partula и Achatinella, уже вымерли, род Amphidromus, охватывающий более 110 видов, является уникальным Полезное для изучения эволюции асимметрии у животных, и именно поэтому Сохранение этого рода имеет важное значение для биологов.

В А. floresianus, подрод Syndromus, скручивание раковины обычно левостороннее. Шкала шкалы 10 мм.

Раковины видов амфидрома А. perversus может быть правым, как показано здесь.

Но скручивание раковины у A. perversus также может быть левым, как показано здесь.

Вид сбоку левосторонней раковины (слева) и вид через отверстие правой раковины (справа) A. perversus
 Amphidromus atricallosus leucoxanthus имеет типичную форму раковины для рода Amphidromus: умеренно выпуклый и гладкий.
Amphidromus atricallosus leucoxanthus имеет типичную форму раковины для рода Amphidromus: умеренно выпуклый и гладкий. Мутовки раковины у видов Amphidromus умеренно выпуклые и за некоторыми исключительными, гладкими или имеют слабую скульптуру роста линий. Amphidromus costifer Смит из провинции Биньдинь во Вьетнаме, смит из провинции Биньдинь во Вьетнаме; Морле из Камбоджи; Таппароне-Канефри из Целебеса; и - комплекс с Явы и Суматры. С ребристостью коррелирует светлая монохромная окраска и тонкая оболочка с большой апертурой и расширяющейся кромкой. Многие твердые раковины у других видов действительно демонстрируют небольшую шероховатость поверхности, но это сильно отличается от упомянутой выше ребристой скульптуры.
Отверстие обычно велико, от двух пятых до одной трети диаметра. высота панциря, часто в пределах одной и той же популяции. Обычно губа хотя бы несколько расширена, и у таких форм, как Schepman и (Pfeiffer) var. inauris Fulton, губу можно назвать только расширяющейся. У (Линней) и других видов с толстой оболочкой губа утолщена изнутри, образуя «валик» при расширении, и имеет очень тяжелую париетальную мозоль. У видов с тонкой оболочкой губа обычно представляет собой простой отраженный край. Область пуповины может быть частично открытой, почти закрытой или герметичной. Эта функция иногда является полезным критерием для идентификации. Угол теменной системы обсуждается, но точной информации об этом не было.
Как правило, обороты раковины увеличиваются в размере довольно регулярно, однако у видов, которые, возможно, связаны между собой, такие как (Рив) и Tapparone-Canefri, могут иметь совершенно разную степень прироста оборотов. Попыток выразить эти различия меристически не предпринималось, поскольку большая часть доступного материала не подходила для статистической обработки. Фактические размеры раковины сильно различаются как внутри, так и между видами. Минимальный размер взрослой особи составляет около 21 мм, максимальный размер - около 75 мм. Размер взрослой особи внутри вида не сильно различается: только несколько видов, в частности, имеют вариацию в размере взрослой особи, которая в целом семь или восемь миллиметров.
 панцирь ярко окрашен, что типично для древесных улиток в целом.
панцирь ярко окрашен, что типично для древесных улиток в целом.  Темный варикоз на раковине Amphidromus perversus отмечает стадию покоя.
Темный варикоз на раковине Amphidromus perversus отмечает стадию покоя. Единственный наиболее важный аспект вариации раковины в пределах рода это цветовой узор. В целом, многие древесные улитки ярко окрашены, очевидными примерами являются роды булимулидов Drymaeus и Liguus, цеполид Polymita и камаениды Папуина. Тем не менее, Polymita, Liguus и Amphidromus особенно известны своими цветовыми вариациями. Основной цвет Amphidromus кажется желтым, и этот цвет обычно ограничен поверхностными слоями раковины, поскольку изношенные экземпляры кажутся почти лишенными основного цвета. У некоторых видов цвет фона беловатый, а у некоторых - темный. апикальные мутовки бледные, пурпурные, коричневые или черные, и это иногда меняется в пределах популяции (как в). Несколько видов, например A. schomburgki, лиственно-зеленый периостракум.
Непрерывные зональные узоры могут принимать форму беловатых субстанций шовных полос (), сильную субпериферическую пигментацию (A. perversus var. Infraviridis), подшивные цветные линии (), широкие спиральные цветные полосы (,) или узкие спиральные полосы (A. laevus ). Прерывистая зональность может состоять из прерывания полос на точки в (); крайне неравномерное разделение зон (vars. sultanus и interruptus); образование наклонных радиальных полос, которые проходят (в A. inversus ) или пересекают (в) линии инкрементного роста; или почти все мыслимые комбинации и вариации этих факторов. Часто паттерн радикально меняется от вершины к витку тела (в вариациях). Апертура, париетальная мозоль, колумелла, губа и пупочная область по-разному отмечены розовым, коричневым, пурпурным, белым или черным. Ханиэль (1921) включает несколько цветных пластинок, которые демонстрируют степень изменения цвета у двух видов типа Syndromus. и подрода Amphidromus одинаково изменчивы, тогда как виды, такие как A. inversus, почти одинаковы по окраске.
В раковинах видов подрода Amphidromus на стадии покоя отмечены отложением коричневая или черная радиальная полоса, называемая варикозом. Это, по-видимому, редко встречается у подрода Syndromus, хотя на панцире A. laevus действительно обнаруживаются прерывания спирального кольцевания после фазы покоя.
Распознавание видов основано на сочетания незначительных структурных изменений, апертуры, контура завитка, области пупка и цветового рисунка. Похоже, что многие виды имеют стабильный цветовой рисунок, в то время как другие виды сильно различаются. Адекватные неотобранные полевые образцы лучше относительной стабильности или изменчивости различных видов в отдельных местах.
Информация о мягкой анатомии Amphidromus широко разрознена и отрывочна. Наиболее полный отчет (1895 г.) об образцах из Большой Натуны (Острова Натуна ) и (Острова Анамба ). К сожалению, хотя, очевидно, что анатомические различия существуют у двух видов, рассеченных Якоби, к сожалению, мы не знаем, какой формой он работал, потому что он неправильно определил свой материал. В своей статье он назвал эти два вида Amphidromus chloris и фазу прерывания A. perversus. Это невозможно, поскольку на самом деле Amphidromus chloris - это вид, встречающийся только на Филиппинских островах, а фаза прерывания A. perversus отсутствует на островах Натуна.
(1893, 1898) обсуждали части анатомии А. adamsii, А. porcellanus, А. Contrarius, и. Уолтер Эдвард Коллиндж (1901, 1902) кратко обозначил особенности А. Palaceus и (сообщается как A. perversus). Ханиэль (1921) проанализировал A. Contrarius и Бернхард Ренш опубликовал несколько разрозненных заметок в своих различных фаунистических обзорах. Несколько более ранних примечаний упоминаются в Пилсбри (1900 г.).
Такие признаки, как длинная узкая почка с загнутым мочеточником и закрытым вторичным мочеточником мочеточником, пениальный комплекс с отчетливым пенисом, непрерывен. с эпифаллосом, эпифаллической слепой кишкой (жгутик и отросток), неразветвленным гаметолитическим протоком, отсутствием вагинальных дополнительных органов и основным состоянием нервной и ретракторной мышечной системы, поддерживающим включение Amphidromus в семейство Camaenidae. Эта группа улиток встречается в самых разных средах обитания в тропиках Восточной Азии и Австралазии, и является одной из самых различных семейства в кладе Стиломматофора. Хотя Laidlaw Solem (1961) не предоставил дополнительных подробностей об анатомии Amphidromus, последующие различных авторов, например, Bishop (1977) и Solem (1983), такие исследования, например, репродуктивная система может предоставить ценные данные для распознавания видов.
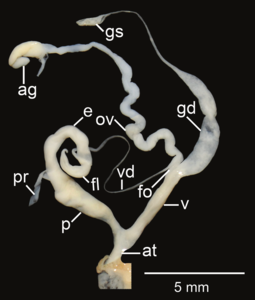
Amphidromus areolatus ; at- предсердие; e– эпифаллус ; fl- жгутик; fo - свободный яйцевод ; gd- проток; gs - гаметолитический мешок; ov– яйцевод ; p- пенис; пр - втягивающая мышца полового члена; v - влагалище; vd– семявыносящий проток
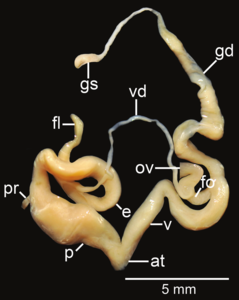
Amphidromus flavus ; at- предсердие; e– эпифаллус ; fl- жгутик; fo - свободный яйцевод ; gd- проток; gs - гаметолитический мешок; ov– яйцевод ; p- пенис; пр - втягивающая мышца полового члена; v - влагалище; vd– семявыносящий проток
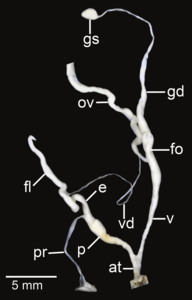
Amphidromus fuscolabris ; at- предсердие; e– эпифаллус ; fl- жгутик; fo - свободный яйцевод ; gd- проток; gs - гаметолитический мешок; ov– яйцевод ; p- пенис; пр - втягивающая мышца полового члена; v - влагалище; vd– семявыносящий проток
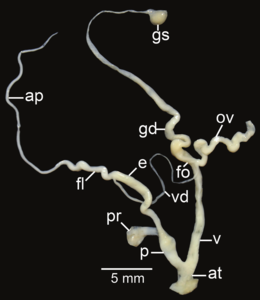
Amphidromus roseolabiatus ; ap- отросток; at - атриум; e– эпифаллус ; fl- жгутик; fo - свободный яйцевод ; gd- проток; gs - гаметолитический мешок; ov– яйцевод ; p- пенис; пр - втягивающая мышца полового члена; v - влагалище; vd– семявыносящий проток
Amphidromus Syndromoideus ; ap- отросток; at - атриум; e– эпифаллус ; fl- жгутик; fo - свободный яйцевод ; gd- проток; gs - гаметолитический мешок; ov– яйцевод ; p- пенис; пр - втягивающая мышца полового члена; v - влагалище; vd– семявыносящий проток

Amphidromus xiengensis ; at- предсердие; e– эпифаллус ; fl- жгутик; fo - свободный яйцевод ; gd- проток; gs - гаметолитический мешок; ov– яйцевод ; p- пенис; пр - втягивающая мышца полового члена; v - влагалище; vd– семявыносящий проток
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Филогения и отношения Amphidromus согласно Sutcharit et al. (2007) |
До 1900 года сходство формы раковины Amphidromus с раковиной южноамериканских древесных улиток семейства Bulimulidae вводило систематиков в заблуждение. Однако вскрытия, сделанные Вигманном и Якоби, ясно показали, что внутренние анатомические особенности Amphidromus были такими же, как и у азиатско-индонезийских Camaenidae, и что сходство раковины Amphidromus с панцирем булимулидов было просто пример параллелизма.
Laidlaw и Solem (1961) распознали 75 видов в роде Amphidromus и поместили еще семь названий в раздел incertae sedis. В 2010 г. было признано 87 видов рода Amphidromus. Виды в пределах рода Amphidromus представлены в следующем списке:
Первоначально описан как Helix perversus Linnaeus in, Amphidromus perversus типовым видом рода Amphidromus, в последующем обозначении Эдуарда фон Мартенса (1860). Виды подрода Amphidromus являются амфидромными (в популяции используются левосторонние и правосторонние улитки) за некоторыми исключениями. Четыре правых таксона: А. Живанши, А. schomburgki dextrochlorus и A. inversus annamiticus; и один левый: А. atricallosus classiaris. Раковины этих видов обычно имеют следующие характеристики: они большие (высота часто превышает 35 мм (1,4 дюйма)); у них соотношение высоты / ширины менее 1,85; и цвет скорлупы желтоватый или зеленоватый. Анатомически у них длинный эпифаллос и жгутик, и обычно присутствует отросток.
Все, кроме двух видов в подроде Syndromus, являющиеся левосторонними. Исключение составляют амфидромин А. glaucolarynx и правый A. круехни. Типовой вид подрода Syndromus - А. Contrarius Müller, 1774, с последующим обозначением (1960). Виды подрода Syndromus имеют раковины меньшего размера (высота обычно менее 35 мм (1,4 дюйма) и соотношение высота / ширина более 1,85) с изменяющимся цветным рисунком. У них также есть короткий эпифаллос и жгутик без отростка. Третий возможный подрод, Goniodromus Bülow, 1905 (типовой вид Amphidromus büllowi Fruhstorfer, 1905), также упоминается в литературе, хотя его подродовой статус еще не подтвержден.
Молекулярный анализ частичных последовательностей 16S рДНК 18 различных видов, проведенные Sutcharit с коллегами (2007), указывают на то, что Amphidromus является монофилетической группой. В их исследовании представлены кладограммы, полученные с помощью различных методов, таких как максимальная экономия, присоединение соседей и максимальная вероятность, были согласованы между собой. Хотя топология , полученная для подрода Amphidromus в соответствии с текущей таксономии, филогения левосторонних видов Syndromus не показала такого соответствия. Кроме того, согласно их результатам, энантиоморфия, по-видимому, является предковым состоянием свертывания раковины в роде Amphidromus, что противоречит общему ожиданию наличия правого свертывания раковин как предкового состояния.
Несмотря на морфологическую идентичность, некоторые экземпляры предположительно принадлежащие к трем видам, а именно Amphidromus semitessellatus, A. xiengensis и A. areolatus, по-видимому, имели полифилетическое происхождение мтДНК гаплотипов. Это привело к тому же одновременно в разных кладах по топологии: например, A. areolatus можно найти в двух разных кладах на кладограмме Сутчарита и коллеги (2007), сгруппированных соответственно с A. xiengiensis, а также с A.semitesselatus. По мнению авторов, эти результаты можно объяснить конвергентными и полиморфными цветными узорами раковины (например, раковины особей имели очень похожие цвета и форму, хотя маркеры мтДНК показали значимые различия). Альтернативно, они также могут быть результатом интрогрессивной гибридизации или предкового полиморфизма мтДНК. В любом случае анализ филогеографии с использованием других маркеров (ядерных маркеров или других маркеров мтДНК) или дополнительных морфологических признаков все равно будет необходим для дальнейшего прояснения этих вопросов.
В настоящее время достоверных ископаемых остатков Amphidromus до появления человека не зарегистрировано. (1932) сообщил о находке нескольких экземпляров, которые в доисторические времена были съедены туземцами на центральном Яве; и несколько лет спустя (van Benthem Jutting, 1937, стр. 92–94) тот же автор сообщил об единственном экземпляре A. дворец с острова Ява. Ни одна из этих записей не предшествовала заселению людей, и поэтому они не проливают света на предчеловеческую историю Amphidromus.
 Мегхалая в Индии - одно из многих мест, где могут быть обитатели Amphidromus.
Мегхалая в Индии - одно из многих мест, где могут быть обитатели Amphidromus.  Aestivating Amphidromus roseolabiatus внутри стерильных ветвей папоротников оленьего рога, Platycerium.
Aestivating Amphidromus roseolabiatus внутри стерильных ветвей папоротников оленьего рога, Platycerium. Пара Amphidromus (Syndromus) sp. Красными стрелками может быть выступающая пстраиля (vsp) вагинального стимулятора, которая может быть стимулирующим органом.
Пара Amphidromus (Syndromus) sp. Красными стрелками может быть выступающая пстраиля (vsp) вагинального стимулятора, которая может быть стимулирующим органом. Род Amphidromus простирается от восточной Индии до Юго-Западной Азии (ограничен на севере Гималаи ), в северной Австралии (ограниченно на восток линией Вебера ). Виды Amphidromus встречаются в следующих местах: с холмов Гаро и холмов Хаси Мегхалаи на северо-востоке Индия ; на всей территории Бирмы, Малайского полуострова, Таиланда, Лаоса, Камбоджи, Вьетнама, Индонезия на восток до Сулавеси, островов Банда, Тимора и (но не на Кераме, Буру, Халмахере, остров Батджан, острова Оби, острова Ару и Кей или архипелаг Талауд и некоторые острова-спутники Целебезии); на юге Филиппин, особенно Минданао и Балабак, Палаван ; и в северной Австралии (представлен исключительно Amphidromusognatus ).
Виды Amphidromus - это древесные наземные улитки. Дополнительная информация о привычках и образе жизни видов Amphidromus практически отсутствует - однако эти улитки обычно собираются Рацион Amphidromus неизвестен, но считается, что Amphidromus atricallosus perakensis питается микроскопическими грибами, лишайники или наземные водоросли.
, несмотря на большое разнообразие внутри этого рода, по состоянию на 2017 год. всей литературе.Исследование (1921) описало откладывание яиц у Amphidromus palaceus var. pura at, Западная Ява. В октябрь 1920 года местные жители с сообщением Явы принесли два «гнезда» с улитками, которые только-только начали откладывать яйца; одна улитка сложила внешние листья молодого бамбукового побега и склеила их в остроконечный корнет. Ствол висел вертикально узким концом вверх, а широкое внизу внизу. Верхняя часть мешка была заполнена яйцами при сборе. Улитка медленно спускалась, вращаясь вокруг своей продольной оси, и откладывала яйца, пока вся полость не была заполнена. Если через трещину в корзине яйца попадали на воздух, они быстро высыхали. Через два дня после поимки кладка яиц закончилась, и улитка закрыла отверстие, закрыв еще несколько листьев. На кладку яиц ушло, вероятно, четыре дня, как в начале наблюдений полость была заполнена наполовину. Второе гнездо из сложного таким же образом листьев манго содержало 234 яйца. Объем яиц в каждом случае превышает размер улитки, что указывает на то, что яйца должны быть инкапсулированы непосредственно перед началом. Капсулы были очень тонкими и быстро сохли на воздухе. Октябрь ознаменовал начало сезона дождей и вероятно, это нормальный период размножения. Ван Бентем Юттинг (1950, стр. 493) сообщил, что яйца A. porcellanus начали вылупляться только через десять дней после кладки. Аналогичные привычки строить гнезда были зарегистрированы и для других видов, но полное исследование жизненного цикла не было опубликовано. Вплоть до 1961 г. не было информации о цикле активности, продолжительности жизни, скорости роста и т. Д.
Schilthuizen et al. (2005) описали пространственное преобразование популяции A. inversus в Малайзии. Schilthuizen et al. (2007) представит, что при спаривании с улитками противоположной хиральности имеет место половой отбор. Это означает, что улитки-левши чаще спариваются с улитками-правшами, чем с улитками той же спирали. Кроме того, существуют анатомические адаптации сперматофора и женской части репродуктивной системы для успеха этих спариваний.
Хищники Среди улиток Amphidromus - барбет с красной короной Megalaima rafflesii и, вероятно, другие птицы. Известно, что азиатские змеи из рода Pareas питаются видами Amphidromus, удаляя мягкие части раковин. Многие раковины Amphidromus были найдены в логове крысы в Малайзии.
Эта статья включает текст из ссылок, являющейся общественным достоянием.
| Викискладе есть медиафайлы, связанные с Amphidromus. |