
Войти
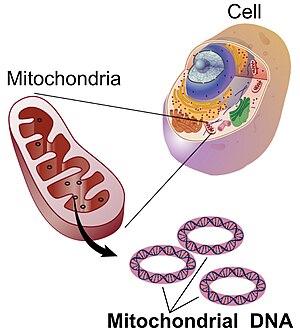 Митохондриальная ДНК - это небольшая кольцевая хромосома, обнаруженная внутри митохондрий. Эти органеллы, обнаруженные в клетках, часто называют электростанциями клетки. Митохондрии и, следовательно, митохондриальная ДНК передаются почти исключительно от матери к потомству через яйцеклетку.
Митохондриальная ДНК - это небольшая кольцевая хромосома, обнаруженная внутри митохондрий. Эти органеллы, обнаруженные в клетках, часто называют электростанциями клетки. Митохондрии и, следовательно, митохондриальная ДНК передаются почти исключительно от матери к потомству через яйцеклетку.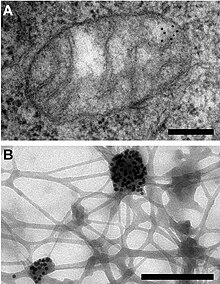 . Электронная микроскопия выявляет митохондриальную ДНК в дискретных очагах. Бары: 200 нм. (A) Цитоплазматический срез после мечения иммунным золотом анти-ДНК; золотые частицы, маркирующие мтДНК, обнаруживаются возле митохондриальной мембраны (черные точки в верхнем углу). (B) Полный вид цитоплазмы после экстракции буфером CSK и мечения иммунным золотом анти-ДНК; мтДНК (отмеченными золотыми частями) сопротивляется экстракции. Из Iborra et al., 2004.
. Электронная микроскопия выявляет митохондриальную ДНК в дискретных очагах. Бары: 200 нм. (A) Цитоплазматический срез после мечения иммунным золотом анти-ДНК; золотые частицы, маркирующие мтДНК, обнаруживаются возле митохондриальной мембраны (черные точки в верхнем углу). (B) Полный вид цитоплазмы после экстракции буфером CSK и мечения иммунным золотом анти-ДНК; мтДНК (отмеченными золотыми частями) сопротивляется экстракции. Из Iborra et al., 2004. Митохондриальная ДНК (мтДНК или мДНК ) - это ДНК, расположенная в митохондриях, клеточные органеллы внутри эукариотических клеток, которые преобразуют химическую энергию из пищи в формулу, клетки могут использовать, аденозинтрифосфат (АТФ). Митохондриальная ДНК - это лишь небольшая часть ДНК в эукариотической клетке; большая часть ДНК может быть обнаружена в ядре клетки, у растений и водорослей также в пластидах, таких как хлоропласты.
Митохондриальная ДНК человека была первая значимая часть генома человека, позвоночная секвенированию. Это секвенирование показало, что мтДНК человека включает в себя 16 569 пар оснований и кодирует быстрее 13 белков.
Механизм развития ядерных генетических маркеров, она представляет собой основу филогенетики и эволюционной биологии. Он также позволяет исследовать родство популяций, что стало важным в антропологии и биогеографии.
Считается, что ядерная и митохондриальная ДНК имеют раздельное эволюционное происхождение, вместе с мтДНК происходящие из кольцевых геномов бактерий, которые были поглощены ранними предками современных эукариотических клеток. Эта теория называется эндосимбиотической теорией. В современных организмах подавляющее большинство существующих в митохондриях современных организмов содержится примерно 1500 различных типов у млекопитающих, кодируется ядерной ДНК, но гены Считается, что если не большинство из них изначально имели бактериальное происхождение, с тех пор они были перенесены в эукариотическое ядро в ходе эволюции.
. Причины, по которым митохондрии сохранили некоторые гены, обсуждаются. Существование у некоторых видов митохондриальных органелл, лишенных генома, предполагает, что возможна полная потеря гена, перенос митохондриальных генов в ядро имеет несколько преимуществ. Сложность доставки удаленно продуцируемых гидрофобных белковых продуктов в митохондрии - одна из гипотез, объясняющих, почему некоторые гены сохраняются в мтДНК; колокализация для окислительно-восстановительной регуляции - другая, вызывающая желательность локального контроля над митохондриальным аппаратом. Недавний анализ широкого спектра генов мтДНК предполагает, что обе эти особенности могут определять удержание митохондриальных генов.
У всех организмов в митохондриальных геномах происходит шесть основных типов генома., классифицируемые по их структуре (например, кольцевой или линейной), размеру, наличию интронов или плазмидоподобных структур, а также по тому, является ли генетический материал единственной молекулы или совокупностью гомогенных или гетерогенные молекулы.
У многих одноклеточных организмов (например, цилиат Tetrahymena и зеленая водоросль Chlamydomonas reinhardtii ), а также в редких случаях у многоклеточных видов (например, у некоторых Cnidaria ) мтДНК обнаруживается как линейно организованная ДНК. Большинство этих линейных мтДНК обладают теломеразой -независимыми теломерами (т.е. концами линейной ДНК ) с различными методами репликации, что сделало их интересными объектами. исследования, потому что многие из этих одноклеточных организмов с линейной мтДНК известны патогенами.
В клетках животных обнаружен только один тип митохондриального генома. Этот геном обычно содержит одну кольцевую молекулу с генетическим языком (тип 1) от 11 до 28 т.п.н. (килограмм пар оснований ).
В феврале 2020 года паразит, связанный с медузами - Henneguya salminicola - было обнаружено, что у него отсутствует митохондриальный геном, но сохраняются структуры, которые считаются связанными с митохондриями органеллами. Более, гены ядерной ДНК, участвующие в аэробном дыхании и репликации и транскрипции митохондриальной ДНК, либо отсутствуют, либо присутствуют только в виде псевдогенов. Это первый многоклеточный организм, у которого отсутствует аэробное дыхание и который живет полностью без кислородной зависимости.
У растений и грибов обнаружены три различных типа митохондриального генома. Первый тип - это кольцевой геном, который имеет интроны (тип 2) и может иметь длину от 19 до 1000 т.п.н. Второй тип генома представляет собой кольцевой геном (около 20–1000 т.п.н.), который также имеет плазмидоподобную устойчивость (1 т.п.н.) (тип 3). Последний тип генома, который можно найти у растений и грибов, - это линейный геном, состоящий из гомогенных молекул ДНК (тип 5).
Существуют большие различия в размере генов мтДНК среди грибов и растений, хотя, по-видимому, существуют основные подмножество генов, которые присутствуют во всех эукариотах (за исключением тех немногих, которые вообще не имеют митохондрий). Некоторые виды растений имеют огромные митохондриальные геномы с мтДНК Silene conica, в составе до 11 300 000 параний. Удивительно, но даже эти огромные мтДНК содержат такие же и виды генов, что и родственные растения с гораздо большим количеством мтДНК. Геном митохондрии огурца (Cucumis sativus ) состоит из трех кольцевых хромосом (длиной 1556, 84 и 45 килобаз), которые полностью или в степени автономны в отношении их репликации.
Протисты содержат самые разные митохондриальные геномы, пять различных типов, встречающихся в этом царстве. Тип 2, тип 3 и тип 5, упомянутые в геномах растений и грибов, также существуют у некоторых протистов, как и два уникальных типа генома. Один из этих уникальных типов представляет собой гетерогенный набор кольцевых молекул ДНК (тип 4), другой - гетерогенный набор линейных молекул (тип 6). Типы генома 4 и 6 имеют размер от 1 до 200 т.п.н.
Наименьший митохондриальный геном, секвенированный на сегодняшний день, - это мтДНК паразита размером 5967 п.н. Plasmodium falciparum.
Эндосимбиотический перенос гена, процесс, посредством которого гены, закодированные в митохондриальном геноме, переносятся в основном геном, вероятно, объясняет, почему более сложные организмы, такие как люди, имеют меньшие митохондриальные геномы, чем более простые организмы, такие как протисты.
| Тип генома | Царство | Интроны | Размер | Форма | Описание |
|---|---|---|---|---|---|
| 1 | Животное | Нет | 11–28 kbp | Круглый | Одномолекулярный |
| 2 | Грибки, растения, протиста | Да | 19–1000 кб | круглая | одиночная молекула |
| 3 | грибы, растения, протиста | № | 20–1000 кбп | круглая | Большие молекулы и малые плазмидоподобные структуры |
| 4 | Protista | № | 1–200 kbp | Circular | Гетерогенная группа молекул |
| 5 | Fungi, Plant, Protista | № | 1–200 kbp | Linear | Однородная группа молекул |
| 6 | Protista | Нет | 1–200 т.п.н. | Линейный | Гетерогенная группа молекул |
Митохондриальная ДНК реплицируется с помощью ДНК-полимеразы гамма-комплекс, состоящий из каталитической ДНК- полимеразы 140 кДа, кодируемой геном POLG и двух аксессуаров 55 кДа убединицы, кодируемые геном POLG2. Реплисомный аппарат образован ДНК-полимеразой, TWINKLE и митохондриальными белками SSB. TWINKLE - это геликаза, которая раскручивает короткие участки дцДНК в направлении от 5 'до 3'. Все эти полипептиды кодируются в ядерном геноме.
Во время эмбриогенеза репликация мтДНК строго подавляется от оплодотворенного ооцита через предимплантационный эмбрион. Результирующее снижение количества копий мтДНК на играет роль в «узком месте» митохондрий, используя межклеточную изменчивость для улучшения наследования повреждающих мутаций. По словам Джастина Сент-Джона и его коллег: «На стадии бластоцисты начало репликации мтДНК специфично для клеток трофэктодермы. Нап, клетки внутренняя клеточная масса ограничивает репликацию мтДНК до тех пор, пока они не получат сигналы на дифференцировать на уровни клеток ».
 Митохондриальная ДНК человека с 37 генами на своих соответствующих H- и L-цепи.
Митохондриальная ДНК человека с 37 генами на своих соответствующих H- и L-цепи. Две цепи митохондриальной ДНК человека различаются как тяжелая цепь и легкая цепь. Тяжелая цепь богата гуанином и кодирует 12 субъединиц системы окислительного фосфорилирования, две рибосомные РНК (12S и 16S) и 14 тРНК. Легкая кодирует одну субъединицу и 8 тРНК. Итак, в целом мтДНК кодирует рРНК, 22 тРНК и 13 белковых субъединиц, все из которых участвуют в процессе окислительного фосфорилирования.
| Ген | Тип | Продукт | Положения. в митогеноме | Нить |
|---|---|---|---|---|
| MT-ATP8 | белок, кодирующий | АТФ-синтазу, субъединица 8 Fo (комплекс V) | 08,366–08,572 (перекрывается с MT-ATP6) | H |
| MT-ATP6 | белок, кодирующий | АТФ-синтазу, субъединицу Fo 6 (комплекс V) | 08,527 –09,207 (перекрывается с MT-ATP8) | H |
| белок MT-CO1 | кодирует | цитохром с оксидазу, субъединица 1 (комплекс IV) | 05,904–07,445 | H |
| MT- CO2 | кодирующий белок | цитохром с оксидазу, субъединица 2 (комплекс IV) | 07,586–08,269 | H |
| MT-CO3 | кодирующий белок | цитохром с оксидазу, субъединица 3 (комплекс IV) | 09,207–09,990 | H |
| кодирующий белок MT-CYB | цитохром b (комплекс III) | 14,747–15,887 | H | |
| кодирование белка MT-ND1 | НАДН-дегидрогеназа, субъединица 1 (комплекс I) | 03,307–04,262 | H | |
| MT-ND2 | белок, кодирующий | НАДН-дегидрогеназу, субъединица 2 (комплекс I) | 04,470–05,511 | H |
| MT-ND3 | белок, кодирующий | НАДН-дегидрогеназу, субъединица 3 (комплекс I) | 10,059–10,404 | H |
| MT-ND4L | белок, кодирующий | НАДН-дегидрогеназу, субъединица 4L (комплекс I) | 10,470–10,766 (перекрывается с MT-ND4) | H |
| MT-ND4 | белок, кодирующий | NADH-дегидрогеназу, субъединица 4 (комплекс I) | 10,760–12,137 (перекрывается с MT-ND4L) | H |
| MT-ND5 | белок, кодирующий | НАДН-дегидрогеназу, субъединица 5 (комплекс I) | 12,337–14,148 | H |
| MT-ND6 | белок, кодирующий | НАДН-дегидрогеназу, субъединица 6 (комплекс I) | 14,149–14,673 | L |
| MT -RNR2 | кодирующий белок | гуманин | — | — |
| MT-TA | транспортная РНК | тРНК- Ала нине (Ala или A) | 05,587–05,655 | L |
| MT-TR | транспортная РНК | тРНК- аргинин (Arg или R) | 10,405–10,469 | H |
| MT-TN | переносящая РНК | тРНК- аспарагин (Asn или N) | 05,657–05,729 | L |
| MT-TD | переносящая РНК | тРНК- аспарагиновая кислота (Asp или D) | 07,518–07,585 | H |
| MT-TC | переносящая РНК | тРНК- Цистеин (Cys или C) | 05,761–05,826 | L |
| MT-TE | транспортная РНК | тРНК- Глутаминовая кислота (Glu или E) | 14,674–14,742 | L |
| MT-TQ | переносящая РНК | тРНК- Глютамин (Gln или Q) | 04,329–04,400 | L |
| MT-TG | переносящая РНК | тРНК- Глицин (Gly или G) | 09,991– 10,058 | H |
| MT-TH | переносимая РНК | тРНК- гистидин (His или H) | 12,138–12,206 | H |
| MT-TI | перенос РНК | тРНК- изолейцин (Иль или I) | 04,263–04,331 | H |
| MT-TL1 | транспортная РНК | тРНК- лейцин (Leu-UUR или L) | 03,230–03,304 | H |
| MT-TL2 | транспортная РНК | тРНК- лейцин (Leu-CUN или L) | 12,266–12,336 | H |
| МТ-ТК | переносимая РНК | тРНК- лизин (Lys или K) | 08,295–08,364 | H |
| МТ-ТМ | перенос РНК | тРНК- метионин (Met или M) | 04,402–04,469 | H |
| MT-TF | транспортная РНК | тРНК- фенилаланин (Phe или F) | 00,577–00,647 | H |
| MT-TP | переносящая РНК | тРНК- пролин (Pro или P) | 15,956–16,023 | L |
| MT-TS1 | транспортная РНК | тРНК- серин (Ser-UCN или S) | 07,446 –07,514 | L |
| MT-TS2 | транспортная РНК | тРНК- Серин (Ser-AGY или S) | 12,207–12,265 | H |
| МТ-ТТ | транспортная РНК | тРНК- Треонин (Thr или T) | 15,888–15,953 | H |
| MT-TW | транспортная РНК | тРНК- триптофа н (Trp или W) | 05,512–05,579 | H |
| MT-TY | транспортная РНК | тРНК- тирозин (Tyr или Y) | 05,826–05,891 | L |
| МТ-ТВ | переносящая РНК | тРНК- валин (Val или V) | 01,602–01,670 | H |
| MT-RNR1 | рибосомная РНК | Малая субъединица: SSU (12S) | 00,648–01,601 | H |
| MT-RNR2 | рибосомная РНК | Большая субъединица : LSU (16S) | 01,671–03,229 | H |
Между большинством (но не всеми) белками-кодирующими областями присутствуют тРНК (см. карту митохондриального генома человека). Во время транскрипции тРНК приобретают характерную L-форму, которая распознается и расщепляется специфическими ферментами. При процессинге митохондриальной РНК отдельные последовательные мРНК, рРНК и тРНК высвобождаются из первичного транскрипта. Таким образом, свернутые тРНК как пунктуации вторичной структуры.
Промоторы для инициации транскрипции тяжелой и легкой цепей расположены в основной некодирующей области мтДНК называется петлей с ущербом, D-петлей. Имеются данные о том, что транскрипция митохондриальных рРНК регулируется промотором 1 тяжелой цепи (HSP1), а транскрипция полицистронных транскриптов, кодирующих белковые субъединицы, регулируется HSP2.
Измерения уровней мтДНК-кодируемых РНК в тканях крупного рогатого скота показали, что существуют большие различия в экспрессии митохондриальных РНК относительно общей тканевой РНК. Среди 12 исследованных тканей самый высокий уровень экспрессии наблюдался в сердце, затем следовали образцы головного мозга и стероидогенной ткани.
Как установлено это действие трофического гормона АКТГ на клетки коры надпочечников, Экспрессия митохондриальных генов может сильно регулироваться внешними факторами, по-видимому, для усиления синтеза митохондриальных белков, необходимых для производства энергии. Интересно, что хотя экспрессия генов, кодирующих белок, стимулировала АКТГ, уровни митохондриальной 16S рРНК не показали значительных изменений.
В большинстве многоклеточных организмов, мтДНК передается по наследству от матери (по материнской линии). Механизмы для этого включают простое разведение (яйцо содержит в среднем 200000 молекул мтДНК, тогда как здоровые человеческие сперматозоиды, как сообщается, содержат в среднем 5 молекул), деградацию мтДНК сперматозоидов в мужских половых путях и оплодотворенная яйцеклетка; и, по крайней мере, у некоторых организмов, неспособность мтДНК сперматозоидов проникнуть в яйцеклетку. Каким бы ни был механизм, этот паттерн наследования мтДНК с одним родителем (монородительское наследование ) встречается среди различных животных, сообществ растений, а также у грибов.
В исключительных случаях человеческие младенцы иногда наследуют мтДНК как от отца, так и от матери, что приводит к мтДНК гетероплазмии.
В половом размножении, митохондрии обычно наследуются исключительно от матери; митохондрии в сперме млекопитающих обычно разрушаются яйцеклеткой после оплодотворения. Кроме того, митохондрии существуют только в хвосте сперматозоидов, который используется для продвижения сперматозоидов, и иногда хвост теряется во время оплодотворения. В 1999 г. Сообщалось, что митохондрии отцовских сперматозоидов (содержит мтДНК) помечены убиквитином, чтобы выбрать для их последующего разрушения внутри эмбриона. Некоторые методы оплодотворения in vitro, в частности инъекции сперматозоидов в ооцит, могут мешать этому.
Тот факт, что митохондриальная ДНК передается по материнской линии, позволяет исследователям генеалогии проследить материнское происхождение в далеком прошлом. (ДНК Y-хромосомы, унаследованная от отца, используется аналогичным образом для определения патрилинейного анамнеза.) Это обычно выполняется на митохондриальной ДНК человека путем секвенирования гипервариабельные контрольные области (HVR1 или HVR2), а иногда и полную молекулу митохондриальной ДНК, как генеалогический ДНК-тест. HVR1, например, состоит примерно из 440 пар оснований. Эти 440 пар оснований сравниваются с теми же регионами других людей (либо конкретных людей, либо субъектов в базе данных) для определения материнской линии. Чаще всего сравнение проводится с исправленной Кембриджской эталонной последовательностью. Vilà et al. опубликовали исследования, отслеживающие материнское происхождение домашних собак от волков. Концепция Митохондриальной Евы основана на том же анализе, пытаясь обнаружить происхождение человечества, отслеживая происхождение во времени.
Можно ожидать, что сущности, подверженные униродительскому наследованию и практически не имеющие рекомбинации, будут подвержены трещотке Мюллера, накоплению вредныхмутаций до тех пор, пока функциональность не станет потерянный. Популяции животных в митохондриях избегают этого процесса развития, известному как узкое место мтДНК. Узкое место использует случайные процессы в клетке для увеличения межклеточной изменчивости мутантной нагрузки по мере развития организма: одна яйцеклетка с некоторой долей мутантной мтДНК, таким образом, производит эмбрион, в котором разные клетки имеют разные мутантные нагрузки. Затем отбор на клеточном уровне может использоваться для удаления тех клеток с большим мутантным мтДНК, что приводит к стабилизации или снижению мутантной нагрузки между поколениями. Механизм, лежащий в узком месте, обсуждается, недавнее математическое и экспериментальное метаисследование алгоритмов случайного разделения мтДНК при делении клетки и случайного оборота молекул мтДНК внутри клетки.
Наследование мужского митохондриальной ДНК было обнаружено у кур Плимут-Рок. Доказательства подтверждают редкие случаи мужского митохондриальной наследования некоторых млекопитающих. В частности, задокументированные используются для мышей, когда митохондрии, унаследованные от самцов, были отторгнуты. Он также был обнаружен у овец и клон крупного рогатого скота. Зарегистрированы редкие случаи мужской митохондриальной наследования у людей. Хотя многие из этих случаев связаны с клонированными эмбрионами или последующим отторжением отцовских митохондрий, другие документируют in vivo наследование и сохранение в лабораторных условиях.
У двустворчатых моллюсков наблюдается двойное однопородное наследование мтДНК. У этих видов самки есть только один тип мтДНК (F), тогда как самцы имеют мтДНК F-типа в своих соматических клетках, а мтДНК M-типа (которая может отличаться на 30%) в зародышевой линии клетки. Унаследованные по отцу митохондрии также зарегистрированы у некоторых насекомых, таких как плодовые мухи, пчелы и периодические цикады.
Известный метод ЭКО, поскольку донорство митохондрий или митохондриальная заместительная терапия (MRT) приводит к получению потомства, содержащего мтДНК от самки-донора и ядерную ДНК от матери и отца. В процедуре переноса веретена ядро яйцеклетки вставляется в цитоплазму яйцеклетки самки-донора, ядро которой было удалено, но все еще содержит мтДНК самки-донора. Затем составное яйцо оплодотворяется спермой мужчины. Процедура используется, когда женщина с генетически дефектными митохондриями желает произвести потомство и потомство со здоровыми митохондриями. Первым известным ребенком, который родился в результате донорства митохондрий, был мальчик, родившийся у иорданской пары в Мексике 6 апреля 2016 года.
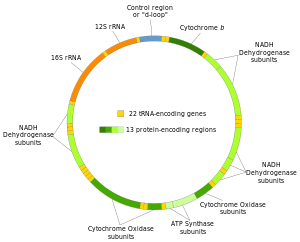 Митохондриальная ДНК человека с группами белков -, гены, кодирующие рРНК и тРНК.
Митохондриальная ДНК человека с группами белков -, гены, кодирующие рРНК и тРНК. 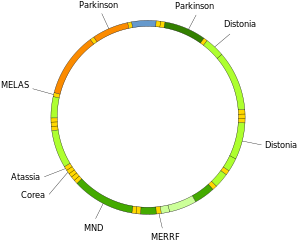 Участие митохондриальной ДНК в некоторых заболеваниях человека.
Участие митохондриальной ДНК в некоторых заболеваниях человека. Концепция, согласно которой мтДНК особенно чувствительна к реактивным формам кислорода генерируется дыхательной цепью из-за ее близости, остается спорным. мтДНК не накапливает больше окислительных повреждений оснований, чем ядерная ДНК. Сообщалось, что по некоторым типам разрушительных повреждений ДНК восстанавливаются более эффективно в митохондриях, чем в ядре. мтДНК упакована с белками, которые кажутся такими же защитными, как белки ядерного хроматина. Более, митохонд развили уникальный механизм, который поддерживает целостность мтДНК за счет деградации чрезмерно поврежденных геномов с помощью репликации интактной / отремонтированной мтДНК. Этот механизм отсутствует в ядре и активируются множественными копиями мтДНК, присутствующими в митохондриях. Результатом мутации мтДНК может быть изменение инструкций по кодированию некоторых белков, что может повлиять на метаболизм и / или приспособленность организма.
Мутации митохондриальной ДНК могут приводить к ряду заболеваний, включая непереносимость физических упражнений и синдром Кернса - Сайра (KSS), который заставляет человека полностью терять функции сердца, глаз и мышц. Некоторые данные свидетельствуют о том, что они могут быть участниками процесса старения и возрастных патологий. В частности, в контексте заболевания доля мутантных молекул мтДНК в клетке называется гетероплазмией. Внутриклеточное и межклеточное распределение гетероплазмии определяет начало и тяжесть заболеваний и находится под сложными стохастическими процессами внутри клетки и во время развития.
Мутации в митохондриальных тРНК могут быть ответственными за тяжелые заболевания, такие как синдромы MELAS и MERRF.
Мутации ядерных генах, кодирующие белки, которые используют митохондрии, также могут быть митохондриальные заболевания. Эти заболевания не следуют паттернам митохондриального наследования, а вместо этого следуют паттернам менделевского наследования.
Недавно мутация в мтДНК была проведена для диагностики рака простаты у пациентов с отрицательным биопсия простаты. Изменения мтДНК могут быть обнаружены в биологических жидкостях больных раком.
Хотя идея свидетельствует о связи между старением и дисфункцией митохондриального генома. По сути, мутации в мтДНК нарушают тщательный баланс производства активных форм кислорода (ROS) и ферментативного ROS (такими ферментами, как супероксиддисмутаза, каталаза, глутатионпероксидаза и другие). Однако некоторые мутации, которые увеличивают выработку АФК (например, за счет снижения антиоксидантной защиты), увеличивают, а не уменьшают их продолжительность жизни. Кроме того, голые землекопы, грызуны размером примерно с мыши живут примерно восемь раз дольше, чем мыши, несмотря на то, что у них снижена по сравнению с мышами антиоксидантная защита и повышена окислительное повреждение биомолекул. Когда-то считалось, что действует цикл положительной обратной связи («порочный цикл»); какохондриальная ДНК накапливает генетические повреждения, вызванные свободными радикалами, митохондрии теряют функцию и утекают свободные радикалы в цитозоль. Снижение митохондриальной функции общую метаболическую эффективность. Эта концепция была окончательно опровергнута, когда были использованы мыши, которые используются генетически для ускоренного накопления мутаций мтДНК, действительно преждевременно стареют, но их не производят больше АФК, как предсказывает гипотеза «порочного цикла». Подтверждая связь между долголетием и митохондриальной ДНК, некоторые исследования корреляции между биохимическими свойствами митохондриальной ДНК и продолжительностью жизни видов. В настоящее время расширенные исследования для дальнейшего изучения этой связи и методов борьбы со старением. В настоящее время генная терапия и нутрицевтики добавки являются популярными областями продолжающихся исследований. Bjelakovic et al. проанализировали результаты 78 исследований с 1977 по 2012 год, в которых участвовало 296 707 участников, и пришли к выводу, что антиоксидантные добавки не снижают общую смертность и не продлевают продолжительность жизни, в то время как некоторые из них, такие как бета- каротин, витамин E и более высокие дозы витамина А, может фактически увеличить смертность.
Повышенное mt повреждение ДНК является признаком нескольких нейродегенеративных заболеваний.
Мозг людей с болезнью Альцгеймера имеют повышенные уровни окислительного повреждения ДНК как в ядерной ДНК, так и в мтДНК, но мтДНК имеет примерно в 10 более высокие уровни, чем ядерная ДНК. Было высказано предположение, что возраст митохондрий является решающим фактором в возникновении нейродегенерации при болезни Альцгеймера.
В болезни Хантингтона мутантный белок хантингтин вызывает дисфункцию митохондрий, включая ингибирование митохондриального транспорта электронов, более высоких уровней активных форм кислорода и повышенный окислительный стресс. Мутантный белок хантингтин способствует окислительному повреждению мтДНК, а также ядерной ДНК, что может вызвать развитие болезни, Гентингтона ология.
продукт окисления ДНК 8-оксогуанин (8-oxoG) - хорошо известный маркер окислительного повреждения ДНК. У людей с боковым амиотрофическим склерозом (БАС) нарушены ферменты, которые обычно восстанавливают повреждения ДНК 8-oxoG в мтДНК спинномозговых мотонейронов. Таким образом, окислительное повреждение мтДНК двигательных нейронов может быть значительным фактором этиологии БАС.
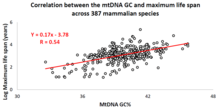 Базовый состав мтДНК видов животных был получен из базы данных MitoAge и сравнен с их максимальной продолжительностью жизни из базы данных AnAge.
Базовый состав мтДНК видов животных был получен из базы данных MitoAge и сравнен с их максимальной продолжительностью жизни из базы данных AnAge. За последнее десятилетие Израильская исследовательская группа под руководством профессора Вадима Фрайфельда показала, что существуют сильные и значимые корреляции между базовым составом мтДНК и максимальной продолжительностью жизни для различных видов животных. Как показано в их работе, высокое содержание мтДНК гуанин + цитозин (GC% ) связано с более длительной максимальной продолжительностью жизни у разных видов животных.. Дополнительное наблюдение включает в том, что корреляция GC% мтДНК с максимальной продолжительностью жизни не зависит от известной корреляции между скоростью метаболизма у животных и максимальной продолжительностью жизни. GC% мтДНК и скорость метаболизма в состоянии покоя объясняют различия в максимальной продолжительности жизни видов животных мультипликативным образом (т.е. максимальная продолжительность жизни = их GC% мтДНК * скорость метаболизма). Чтобы помочь сообществу сообщества в проведении сравнительного анализа характеристик мтДНК и продолжительности жизни животных, была создана специальная база данных под названием MitoAge.
Удаление точки останова часто встречаются внутри или рядом с областями, показывающими неканонические (не B) конформации, такие шпильки, крестообразные элементы и элементы, похожие на клеверный лист. Более того, есть данные, подтверждающие неверные спираль искривленных участковных G-тетрад в возникновении событий нестабильности. Кроме того, наблюдаются более высокие плотности точек разрыва постоянно наблюдаются в областях с перекосом GC и в непосредственной связи от мотива вырожденной последовательности YMMYMNNMMHM.
В отличие от ядерной ДНК, которая унаследована от обоих родителей, и в гены реорганизованы в процессе рекомбинации, обычно нет изменений в мтДНК от родителя к потомству. Хотя мтДНК также рекомбинирует, она делает это с собственными копиями в одной и той же митохондрии. Из-за этого, а также из-за того, что частота мутаций мтДНК животных выше, чем у ядерной ДНК, мтДНК является мощным инструментом для производства по самкам (матрилинейный возраст ) и использовалась роль для установки многих видов наций поколений.
Высокая скорость мутаций (у животных) делает мтДНК полезной для оценки генетических взаимоотношений отдельных лиц или групп внутри вида, а также для идентификации и количественной оценки филогенеза (эволюционных взаимоотношений; см. филогенетика ) разные виды. Для этого биологи определяют и затем сравнивают мтДНК у разных людей или видов. Данные используются для построения сетей взаимодействия между последовательностями, которые обеспечивают взаимодействие между людьми или видами, от которых взяты взяты мтДНК. мтДНК можно использовать для обработки взаимоотношений как между близкородственными, так и между отдаленно родственными видами. Из-за высокой скорости мутаций мтДНК у животных 3-е положения кодонов меняются относительно быстро и, таким образом, предоставляют информацию о генетических расстояниях между близкородственными особями или видами. С другой стороны, скорость замены mt-белков очень низкая, поэтому аминокислотные изменения накапливаются медленно (с соответствующими медленными изменениями в положениях 1-го и 2-го кодона), и, таким образом, они предоставляют информацию о генетических расстояниях отдаленно родственных видов. Статистические модели, которые рассматривают частоту замен между позициями кодонов отдельно, могут, таким образом, использоваться для одновременной оценки филогении, которые содержат как близкородственные, так и отдаленные виды
Митохондриальная ДНК впервые была допущена в качестве доказательства в зале суда США в 1996 г. во время рассмотрения дела «Штат Теннесси против Пола Уэра».
В судебном деле Содружества Пенсильвании против Патриции Линн Роррер в 1998 г. митохондриальная ДНК была впервые допущена в качестве доказательства в штате Пенсильвания. Это дело было показано в 55-й серии 5-го сезона драматического сериала Судебно-медицинская экспертиза (5-й сезон).
Митохондриальная ДНК впервые была допущена в качестве доказательства в Калифорнии, США, в успешном судебное преследование Дэвида Вестерфилда за похищение и убийство 7-летней Даниэль ван Дам в 2002 году в Сан-Диего : оно использовалось для идентификации как человека, так и собаки. Это было первое испытание в США, в котором была допущена собачья ДНК.
Останки короля Ричарда III были идентифицированы путем сравнения его мтДНК с таковыми двух матрилинейных потомков его сестры.
мтДНК сохраняется во всем эукариотическом организме, учитывая критическую роль митохондрий в клеточном дыхании. Однако из-за менее эффективной репарации ДНК (по сравнению с ядерной ДНК) она имеет относительно высокую скорость мутаций (но медленную по сравнению с другими участками ДНК, такими как микросателлиты ), что делает его полезным для изучения эволюционных взаимоотношений - филогения - организмов. Биологи могут определять и затем сравнивать последовательности мтДНК у разных видов и использовать сравнения для построения эволюционного дерева для исследуемого вида.
Например, большинство ядерных генов почти идентичны у людей и шимпанзе, митохондриальные геномы отличаются на 9,8%. Митохондриальные геномы человека и гориллы различаются на 11,8%, что показывает, что мы более связаны с шимпанзе, чем с гориллами.
Митохондриальная ДНК была открыта в 1960-х годах Маргит. М.К. Насс и Сильван Насс с помощью электронной микроскопии в качестве чувствительных к ДНКаз нитей внутри митохондрий, а также по данным Эллен Хаслбруннер, Ханс Таппи и Готфрид Шатц с помощью биохимических анализов на высокоочищенных митохондриальных фракции.
Было создано несколько баз данных для сбора последовательностей митохондриального генома и другой информации. Хотя большинство из них сосредоточено на данных о последовательностях, некоторые из них включают филогенетическую или функциональную информацию.
Существуют несколько стандартных данных, в которых сообщается о полимизмах и мутациях в митохондриальной ДНК человека, а также об оценке их патогенности.