
Войти
Соматическая гипермутация (или SHM ) - это клеточный механизм, с помощью которого иммунная система приспосабливается к новым посторонним элементам, которые ему противостоят (например, микробы ), как видно во время переключения классов. Главный компонент процесса созревания аффинности, SHM диверсифицирует рецепторы В-клеток, используемые для распознавания чужеродных элементов (антигены ), и позволяет иммунной системе адаптировать свой ответ новым угрозам в течение жизни организма. Соматическая гипермутация включает запрограммированный процесс мутации, затрагивающий константные области генов иммуноглобулина. В отличие от мутации зародышевой линии, SHM влияет только на отдельные иммунные клетки организма, и мутации не передаются потомству организма. Неверно направленная соматическая гипермутация - вероятный механизм развития В-клеточных лимфом и многих других видов рака.
Когда В-клетка распознает антиген, она стимулируется к делению ( или размножаться ). Во время пролиферации локус В-клеточного рецептора претерпевает чрезвычайно высокую скорость соматических мутаций, которая по крайней мере в 10–10 раз больше чем нормальная скорость мутации в геноме. Вариации в основном имеют форму одноосновных замен, причем вставки и делеции встречаются реже. Эти мутации происходят в основном в «горячих точках» ДНК, которые сосредоточены в гипервариабельных областях. Эти области соответствуют определяющим комплементарность областям ; сайты, участвующие в распознавании антигена иммуноглобулина. «Горячие точки» соматической гипермутации различаются в зависимости от основания, которое подвергается мутации. RGYW для G, WRCY для C, WA для A и TW для T. Общий результат процесса гипермутации достигается за счет баланса между подверженным ошибкам и высокоточным ремонтом. Эта направленная гипермутация позволяет отобрать В-клетки, которые экспрессируют рецепторы иммуноглобулинов, обладающие повышенной способностью распознавать и связывать специфический чужеродный антиген.
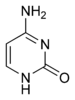 Цитозин
Цитозин  Урацил
Урацил Экспериментальные данные подтверждают точку зрения, что механизм SHM включает дезаминирование цитозина до урацила в ДНК ферментом, называемым цитидиндезаминазой, индуцированной активацией, или AID. Таким образом, пара цитозин: гуанин непосредственно мутирует в несовпадение урацил: гуанин. Остатки урацила обычно не обнаруживаются в ДНК, поэтому для поддержания целостности генома большинство этих мутаций необходимо репарировать с помощью высокоточных ферментов эксцизионной репарации оснований . Основания урацила удаляются ферментом репарации, урацил-ДНК-гликозилазой. Затем привлекаются склонные к ошибкам ДНК-полимеразы, чтобы заполнить пробел и создать мутации.
Синтез этой новой ДНК включает подверженные ошибкам ДНК-полимеразы, которые часто вносят мутации в положение сам дезаминированный цитозин или соседние пары оснований. Во время деления В-клеток ДНК иммуноглобулина вариабельной области транскрибируется и транслируется. Введение мутаций в быстро размножающуюся популяцию В-клеток в конечном итоге приводит к производству тысяч В-клеток, обладающих немного разными рецепторами и различной специфичностью к антигену, из которых В-клетки с наивысшим сродством к можно выбрать антиген. Затем будут отобраны В-клетки с наибольшим сродством для дифференцировки в плазматические клетки, продуцирующие антитело, и долгоживущие В-клетки памяти, способствующие усилению иммунных ответов при повторном заражении..
В процессе гипермутации также используются клетки, которые автоматически выбираются по «сигнатуре» собственных клеток организма. Предполагается, что сбои в этом процессе автоматического выбора могут также привести к развитию аутоиммунного ответа.
Развитие жизнеспособности двух основные конкурирующие молекулярные модели механизма соматической гипермутации (SHM) с 1987 года достигли разрешения, конкретные молекулярные данные опубликованы с 2000 года. Большая часть этих данных ранней фазы была рассмотрена Тенгом и Папавасилиу и дополнительно выделена Ди Нойей и Молом, и полевые данные SHM, рассмотренные в Стиле и дополнительно изложенные в этих статьях.
Это можно назвать моделью на основе ДНК. Он ферментативно ориентирован исключительно на субстраты ДНК. Современная форма, описанная в предыдущих разделах, - это «модель дезаминирования ДНК» Нойбергера, основанная на индуцированной активацией цитидиндезаминазе (AID) и подверженной ошибкам репарации ДНК с помощью ДНК-полимеразы-eta, действующей вокруг поражений AID C-to-U. Модель лишь частично объясняет происхождение полного спектра соматических мутаций в парах оснований A: T и G: C, наблюдаемых в SHM в B-лимфоцитах in vivo во время антиген-управляемого иммунного ответа. Это также не объясняет логически, как могут возникать смещенные по цепи мутации. Ключевой особенностью является его критическая зависимость от свойств синтеза ДНК-полимеразы-эта, нацеленной на пары оснований A: T в AID-опосредованных C-to-U поражениях или разрывах оцДНК, склонных к ошибкам заполнения пропусков, способных к репарации ДНК. Эта подверженная ошибкам ДНК-полимераза является единственной известной подверженной ошибкам полимеразой, участвующей в SHM in vivo. В этих исследованиях часто игнорируется то, что этот фермент ДНК-полимераза семейства Y также является эффективной обратной транскриптазой, что продемонстрировано в анализах in vitro.
 Аденозин
Аденозин  Инозин
Инозин Чем больше Спорный конкурирующий механизм - это основанный на РНК / ОТ механизм (модель обратной транскриптазы SHM), который пытается объяснить получение полного спектра смещенных цепью мутаций в парах оснований A: T и G: C, посредством которых наблюдаются мутации A, превышают мутации T (A>>>T), а мутации G превышают мутации C (G>>>C). Это включает подверженный ошибкам синтез кДНК через РНК-зависимую ДНК-полимеразу, копирующую матрицу пре-мРНК Ig с модифицированным основанием и интегрирующую теперь уже заполненную ошибками копию кДНК обратно в нормальный участок хромосомы. Ошибки в пре-мРНК Ig представляют собой комбинацию редактирования аденозин-инозиновой (A-to-I) РНК и комплекса элонгации транскрипции РНК-полимеразы II, копируя урацил и абазические сайты (возникающие как AID-опосредованные поражения) в зарождающиеся преждевременные клетки. -мРНК с использованием транскрибированной (TS) ДНК в качестве нити копирующей матрицы. Таким образом, современная форма этого механизма в значительной степени зависит от повреждений ДНК AID C-to-U и подверженного ошибкам синтеза кДНК транскрибируемой цепи ДНК-полимеразой-эта, действующей как обратная транскриптаза.
Доказательства того, что и против каждого механизма критически оценен Стил, показывающий, что все молекулярные данные о SHM, опубликованные с 1980 года, прямо или косвенно подтверждают этот основанный на РНК / ОТ механизм. Недавно Zheng et al. предоставили важную независимую проверку, показав, что ферменты аденозиндезаминазы, действующие на РНК (ADAR), могут редактировать как РНК, так и фрагменты ДНК гибридов РНК: ДНК в биохимических анализах in vitro. РНК: гибриды ДНК длиной около 11 нуклеотидов представляют собой временные структуры, образующиеся в пузырьках транскрипции in vivo во время элонгации РНК-полимеразы II.
Предварительный анализ последствий Zheng et al. данные были представлены Стилом и Линдли в виде официального документа в рецензируемый журнал. Zheng et al. данные строго подразумевают, что РНК-фрагмент необходимо сначала отредактировать РНК от A к I, а затем подвергнуть обратной транскрипции и интегрировать, чтобы получить сильные сигнатуры смещенных по цепи A>>>T мутаций в парах оснований A: T, наблюдаемых во всех SHM и гипермутации рака наборы данных. Редактирование (A-to-I) фрагмента ДНК в РНК: ДНК-гибриды in vivo не могут объяснить смещение цепи A>>T, поскольку такие прямые модификации ДНК могут привести к смещению цепи T>>>A, которое не наблюдается ни в одном SHM или набор данных рака in vivo. В этом отношении Робин Линдли также недавно обнаружил, что Ig-SHM-подобные цепочечные мутации в генах, кодирующих белок генома рака, также находятся в «кодонном контексте». Линдли назвал этот процесс направленной соматической мутацией (TSM), чтобы подчеркнуть, что соматические мутации гораздо более целенаправленны, чем считалось ранее, в соматических тканях, связанных с заболеванием. Процесс TSM подразумевает «считыватель ДНК в рамке», посредством которого ДНК и РНК-дезаминазы в транскрибируемых областях управляются в их мутагенном действии с помощью рамки считывания кодонов ДНК.