
Войти
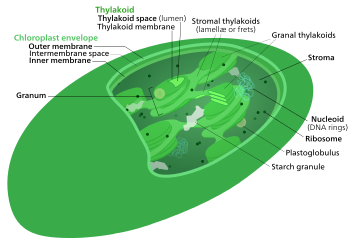 Внутренняя структура хлоропласта
Внутренняя структура хлоропласта Цикл Кальвина, светонезависимые реакции, биосинтетическая фаза, темновые реакции или цикл фотосинтетического восстановления углерода (ПЦР) фотосинтеза - химические реакции, в результате которых диоксид углерода и другие соединения преобразуются в глюкозу. Эти реакции происходят в строме, заполненной жидкостью области хлоропласта за пределами тилакоидных мембран. Эти реакции берут продукты (АТФ и НАДФ ) светозависимых реакций и проводят с ними дальнейшие химические процессы. [Цикл Кальвина использует восстанавливающие силы АТФ и НАДФН из светозависимых реакций для производства сахаров для растений. Эти субстраты используются в серии окислительно-восстановительных реакций для получения сахаров ступенчатым способом. Не существует прямой реакции, которая превращает СО2 в сахар, потому что вся энергия будет потеряна на тепло.] Есть три фазы светонезависимых реакций, которые вместе называются циклом Кальвина: фиксация углерода, реакции восстановления и рибулоза 1, Регенерация 5-бисфосфата (RuBP).
Хотя это и называется «реакцией темноты», цикл Кальвина на самом деле не происходит в темноте или в ночное время. Это связано с тем, что для этого процесса требуется пониженный уровень НАДФ, который является недолговечным и возникает в результате светозависимых реакций. В темноте растения вместо этого выделяют сахарозу во флоэму из своих запасов крахмала, чтобы обеспечить растение энергией. Таким образом, цикл Кальвина происходит, когда свет доступен независимо от типа фотосинтеза (фиксация углерода C3, фиксация углерода C4 и метаболизм крассулоидной кислоты (CAM) ) ; Растения CAM хранят яблочную кислоту в своих вакуолях каждую ночь и высвобождают ее днем, чтобы заставить этот процесс работать.
Эти реакции тесно связаны в тилакоидную цепь переноса электронов, поскольку энергия, необходимая для восстановления диоксида углерода, обеспечивается НАДФН, образующимся в фотосистеме I во время светозависимых реакций. Процесс фотодыхания, также известный как цикл C2, также связан с циклом Кальвина, поскольку он является результатом альтернативной реакции фермента RuBisCO, и его конечным побочным продуктом является другой глицеральдегид. -3-П.
 Обзор цикла Кальвина и фиксации углерода
Обзор цикла Кальвина и фиксации углерода цикл Кальвина, цикл Кальвина – Бенсона – Бассама (CBB), восстановительный пентозофосфатный цикл (RPP-цикл) или C3-цикл представляет собой серию биохимических окислительно-восстановительных реакций, которые происходят в строма хлоропласта в фотосинтетических организмах.
Цикл был открыт в 1950 году Мелвином Кэлвином, Джеймсом Бэшем и Эндрю Бенсон из Калифорнийского университета в Беркли с использованием радиоактивного изотопа углерода-14.
Фотосинтез в клетке происходит в два этапа. На первом этапе светозависимые реакции улавливают энергию света и используют ее для создания молекул хранения и транспорта энергии АТФ и НАДФН. Цикл Кальвина использует энергию короткоживущих электронно-возбужденных носителей для преобразования двуокиси углерода и воды в органические соединения, которые могут быть использованы организмом (и животные, которые им питаются). Этот набор реакций также называется фиксацией углерода. Ключевой фермент цикла называется RuBisCO. В следующих биохимических уравнениях химические соединения (фосфаты и карбоновые кислоты) находятся в равновесии между своими различными ионизированными состояниями, что регулируется pH.
. Ферменты цикла Кальвина функционально эквивалентны большинству ферментов, используемых в других метаболических путях, таких как глюконеогенез и пентозофосфатный путь, но они обнаруживаются в строме хлоропласта вместо клеточного цитозоля, разделяя реакции. Они активируются на свету (поэтому название «темная реакция» вводит в заблуждение), а также продуктами светозависимой реакции. Эти регулирующие функции не позволяют циклу Кальвина вдыхать двуокись углерода. Энергия (в форме АТФ) будет потрачена на проведение этих реакций, которые не имеют чистой производительности.
Сумма реакций в цикле Кальвина следующая:
Гексозные (шестиуглеродные) сахара не являются продуктом цикла Кальвина. Хотя во многих текстах продукт фотосинтеза обозначен как C. 6H. 12O. 6, в основном это удобство для противодействия уравнению дыхания, в котором шестиуглеродные сахара окисляются в митохондриях. Углеводные продукты цикла Кальвина представляют собой трехуглеродные молекулы фосфата сахара или «триозофосфаты», а именно глицеральдегид-3-фосфат (G3P).
На первой стадии цикла Кальвина молекула CO. 2 включается в одну из двух трехуглеродных молекул (глицеральдегид-3-фосфат или G3P), где израсходованы две молекулы АТФ и две молекулы NADPH, которые были произведены на светозависимой стадии. Три задействованных этапа:
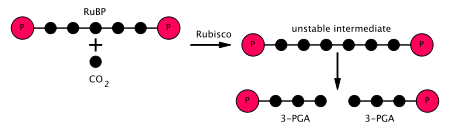 этап 1 цикла Кальвина (черные кружки представляют атомы углерода)
этап 1 цикла Кальвина (черные кружки представляют атомы углерода)  объединение этапов 2 и 3 цикла Кальвина
объединение этапов 2 и 3 цикла Кальвина  Этап регенерации цикла Кальвина
Этап регенерации цикла Кальвина Следующим этапом цикла Кальвина является регенерация RuBP. Пять молекул G3P производят три молекулы RuBP, используя до трех молекул АТФ. Поскольку каждая молекула CO. 2 производит две молекулы G3P, три молекулы CO. 2 производят шесть молекул G3P, пять из которых используются для регенерации RuBP, в результате чего получается чистая прибыль в размере одной молекулы G3P на три молекулы CO. 2. (как и следовало ожидать от количества задействованных атомов углерода).
 Упрощенный цикл C3 со структурными формулами
Упрощенный цикл C3 со структурными формулами Этап регенерации можно разбить на этапы.
Таким образом, из шести продуцируемых G3P пять используются для образования трех молекул RuBP (5C) (всего 15 атомов углерода), и только один G3P доступен для последующего преобразования в гексозу. Для этого требуется девять молекул АТФ и шесть молекул НАДФН на три молекулы СО. 2. Уравнение всего цикла Кальвина схематично показано ниже.
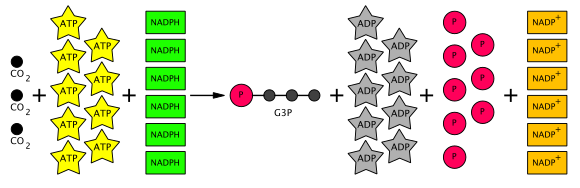 Общее уравнение цикла Кальвина (черные кружки представляют атомы углерода)
Общее уравнение цикла Кальвина (черные кружки представляют атомы углерода) RuBisCO также конкурентно реагирует с O. 2вместо CO. 2в фотодыхании. Скорость фотодыхания выше при высоких температурах. Фотодыхание превращает RuBP в 3-PGA и 2-фосфогликолят, молекулу с 2 атомами углерода, которая может быть преобразована через гликолат и глиоксалат в глицин. Через систему расщепления глицином и тетрагидрофолат два глицина превращаются в серин + CO. 2. Серин можно снова превратить в 3-фосфоглицерат. Таким образом, только 3 из 4 атомов углерода из двух фосфогликолатов могут быть преобразованы обратно в 3-PGA. Можно видеть, что фотодыхание имеет очень негативные последствия для растений, потому что вместо фиксации CO. 2этот процесс приводит к потере CO. 2. . Фиксация углерода C4, разработанная для предотвращения фотодыхания, может происходить только в некоторые растения, произрастающие в очень теплом или тропическом климате, например кукуруза.
Непосредственными продуктами одного витка цикла Кальвина являются 2 молекулы глицеральдегид-3-фосфата (G3P), 3 АДФ и 2 НАДФ. (АДФ и НАДФ на самом деле не являются «продуктами». Они регенерируются и позже снова используются в светозависимых реакциях ). Каждая молекула G3P состоит из 3 атомов углерода. Для продолжения цикла Кальвина необходимо регенерировать RuBP (рибулозо-1,5-бисфосфат). Таким образом, для этой цели используются 5 из 6 атомов углерода в 2 молекулах G3P. Следовательно, для игры за каждый ход производится только 1 чистый углерод. Чтобы создать 1 избыток G3P, требуется 3 атома углерода и, следовательно, 3 витка цикла Кальвина. Чтобы создать одну молекулу глюкозы (которую можно создать из 2 молекул G3P), потребуется 6 витков цикла Кальвина. Избыток G3P также может быть использован для образования других углеводов, таких как крахмал, сахароза и целлюлоза, в зависимости от того, что нужно растению.
Эти реакции не происходят в темноте или ночью. Существует светозависимая регуляция ферментов цикла, так как третья стадия требует пониженного НАДФ.
. Когда цикл должен быть включен или выключен, действуют две системы регулирования: тиоредоксин / система активации ферредоксина, которая активирует некоторые ферменты цикла; и активация фермента RuBisCo, активного в цикле Кальвина, который включает его собственную активазу.
Система тиоредоксин / ферредоксин активирует ферменты глицеральдегид-3-P дегидрогеназу, глицеральдегид-3-P фосфатазу, фруктозо-1,6-бисфосфатазу, седогептулозо-1,7-бисфосфатазу и рибулозо-5-фосфатазу. киназы, которые являются ключевыми точками процесса. Это происходит, когда доступен свет, поскольку белок ферредоксин восстанавливается в комплексе фотосистемы I тилакоидной электронной цепи, когда электроны циркулируют через него. Затем ферредоксин связывается с белком тиоредоксина и восстанавливает его, который активирует ферменты цикла, разрывая связь цистина, обнаруженную во всех этих ферментах. Это динамический процесс, поскольку такая же связь снова образуется другими белками, которые дезактивируют ферменты. Последствия этого процесса заключаются в том, что ферменты в основном остаются активированными днем и дезактивируются в темноте, когда восстановленный ферредоксин больше не доступен.
Фермент RuBisCo имеет собственный, более сложный процесс активации. Это требует, чтобы конкретная аминокислота лизин была карбамилирована для активации фермента. Этот лизин связывается с RuBP и приводит к нефункциональному состоянию, если его оставить некарбамилированным. Специфический фермент активаза, называемый RuBisCo activase, помогает процессу карбамилирования, удаляя один протон из лизина и делая возможным связывание молекулы диоксида углерода. Но даже в этом случае фермент RuBisCo еще не работает, так как для функционирования ему необходим ион магния, связанный с лизином. Этот ион магния высвобождается из просвета тилакоида, когда внутренний pH падает из-за активной откачки протонов из потока электронов. Сама активаза RuBisCo активируется повышенными концентрациями АТФ в строме, вызванными его фосфорилированием.