
Войти
| 5S рибосомная РНК | |
|---|---|
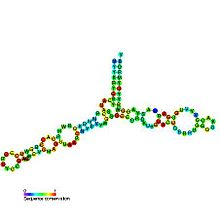 Предсказанная вторичная структура и сохранение последовательности 5S рибосомной РНК Предсказанная вторичная структура и сохранение последовательности 5S рибосомной РНК | |
| Идентификаторы | |
| Символ | 5S_rRNA |
| Rfam | RF00001 CL00113 |
| Прочие данные | |
| РНК тип | Ген ; рРНК |
| Домен (ы) | Eukaryota ; Бактерии ; Археи |
| GO | 0005840 0003735 |
| SO | 0000652 |
| PDB структуры | PDBe |
5S рибосомная РНК (5S рРНК ) представляет собой молекулу рибосомной РНК длиной приблизительно 120 нуклеотидов с массой 40 кДа. Это структурный и функциональный компонент большой субъединицы рибосомы во всех сферах жизни (бактерии, археи и эукариоты ), за исключением митохондриальных рибосом грибов и животных. Обозначение 5S относится к скорости седиментации молекулы в ультрацентрифуге, которая измеряется в единицах Сведберга (S).
 Рисунок 1: Трехмерное представление 5S молекула рРНК. Эта структура представляет собой 5S рРНК из 50S субъединицы рибосомы Escherichia coli и основана на криоэлектронно-микроскопической реконструкции.
Рисунок 1: Трехмерное представление 5S молекула рРНК. Эта структура представляет собой 5S рРНК из 50S субъединицы рибосомы Escherichia coli и основана на криоэлектронно-микроскопической реконструкции.У прокариот ген 5S рРНК обычно расположен в оперонах рРНК ниже малых и большая субъединица рРНК и котранскрибируется в полицистронный предшественник. Особенностью ядерных геномов эукариот является наличие множественных копий гена 5S рРНК (5S рДНК), сгруппированных в тандемные повторы, причем число копий варьируется от вида к виду. 5S рРНК эукариот синтезируется РНК-полимеразой III, в то время как другие эукариотические рРНК отщепляются от предшественника 45S, транскрибируемого РНК-полимеразой I. В ооцитах Xenopus было показано, что пальцы 4-7 девяти- цинкового пальца фактора транскрипции TFIIIA могут связываться с центральная область 5S РНК. Связывание между 5S рРНК и TFIIIA служит как для подавления дальнейшей транскрипции гена 5S РНК , так и для стабилизации транскрипта 5S РНК до тех пор, пока он не потребуется для сборки рибосомы.
вторичная структура 5S рРНК состоит из пяти спиралей (обозначенных IV в римскими цифрами ), четырех петель (BE) и одного шарнира (A), который вместе образуют Y-образную структуру. Петли C и D - концевые шпильки, а петли B и E - внутренние. Согласно филогенетическим исследованиям, спирали I и III, вероятно, являются наследственными. Helix III включает два высококонсервативных аденозина. Считается, что спираль V со своей шпилечной структурой взаимодействует с TFIIIA.
 Рисунок 2: Атомная трехмерная структура 50S-субъединицы из Haloarcula marismortui, ПДБ 1ФФК. Белки показаны синим, 23S рРНК - оранжевым, а 5S рРНК - желтым. 5S рРНК вместе с рибосомными белками L5 и L18 и доменом V 23S рРНК составляют основную часть центрального выступа (ЦП).
Рисунок 2: Атомная трехмерная структура 50S-субъединицы из Haloarcula marismortui, ПДБ 1ФФК. Белки показаны синим, 23S рРНК - оранжевым, а 5S рРНК - желтым. 5S рРНК вместе с рибосомными белками L5 и L18 и доменом V 23S рРНК составляют основную часть центрального выступа (ЦП). Использование различных молекулярных методов, включая иммуно-электронную микроскопию, криоэлектронная микроскопия, межмолекулярное химическое сшивание и рентгеновская кристаллография, было установлено, что расположение 5S рРНК внутри большой рибосомной субъединицы большая точность. У бактерий и архей большая рибосомная субъединица (LSU) сама по себе состоит из двух частей РНК, 5S рРНК и другой более крупной РНК, известной как 23S рРНК, наряду с многочисленными ассоциированными белками.
У эукариот LSU содержит 5S, 5,8S и 28S рРНК и даже больше белков. Структура LSU в 3-х измерениях показывает одну относительно гладкую поверхность и противоположную поверхность, имеющую три выступа, а именно выступ L1, центральный выступ (CP) и ножку L7 / L12. Выступ L1 и ножка L7 / L12 расположены латерально вокруг ЦП. 5S рРНК расположена в ЦП и участвует в формировании и структуре этого выступа. Другие основные составляющие центрального выступа включают 23S рРНК (или, альтернативно, 28S у эукариот) и несколько белков, включая L5, L18, L25 и L27.
Точная функция 5S рРНК еще не ясна. В Escherichia coli делеции гена 5S рРНК снижают скорость синтеза белка и оказывают более сильное пагубное влияние на приспособленность клеток, чем делеции сопоставимого количества копий другого (16S и 23S ) гены рРНК. Кристаллографические исследования показывают, что 5S рРНК-связывающие белки и другие белки центрального выступа LSU играют роль в связывании тРНК. Кроме того, топографическая и физическая близость между 5S рРНК и 23S рРНК, которая формирует пептидилтрансферазу и центр, связывающий GTPase, предполагает, что 5S рРНК действует как посредник между двумя функциональными центрами рибосомы, образуя вместе с 5S рРНК-связывающими белки и другие компоненты центрального выступа, межсубъединичных мостиков и сайтов связывания тРНК.
У эукариот цитозольная рибосома собирается из четырех рРНК и более 80 белков. После транскрибирования 3'-концы 5S рРНК процессируются экзонуклеазами Rex1p, Rex2p и Rex3p. Субъединицы рибосомы 60S и 40S экспортируются из ядра в цитоплазму, где они соединяются с образованием зрелой и трансляционной -компетентной 80S рибосомы. Вопрос о том, когда именно 5S рРНК интегрирован в рибосому, остается спорным, но обычно считается, что 5S рРНК включается в частицу 90S, которая является предшественником частицы 60S, как часть небольшого независимого от рибосомы комплекса РНП, образованного 5S рРНК и рибосомный белок L5.
Несколько важных белков, которые взаимодействуют с 5S рРНК, перечислены ниже.
Взаимодействие 5S рРНК с белком La предотвращает расщепление РНК экзонуклеазами в клетке. белок La обнаружен в ядро у всех эукариотических организмов и ассоциирует с несколькими типами РНК, транскрибируемых РНК pol III. Белок La взаимодействует с этими РНК (включая 5S рРНК) через их 3'-олиго-уридиновый тракт, способствуя стабильности и укладке РНК.
В эукариотических клетках рибосомный белок L5 связывает и стабилизирует 5S рРНК, образуя прерибосомную рибонуклеопротеиновую частицу (RNP), которая находится как в цитозоле, так и в ядре. Дефицит L5 предотвращает транспорт 5S рРНК в ядро и приводит к снижению сборки рибосом.
У прокариот 5S рРНК связывается с рибосомными белками L5, L18 и L25, тогда как у прокариот Известно, что 5S рРНК эукариот связывается только с рибосомным белком L5. У T. brucei, возбудителя сонной болезни, 5S рРНК взаимодействует с двумя близкородственными РНК-связывающими белками, P34 и P37, потеря которых приводит к более низкому глобальному уровню 5S рРНК.
| пермутированный митохондриальный геном, кодируемый 5S рРНК | |
|---|---|
| Идентификаторы | |
| Символ | mtPerm-5S |
| Rfam | RF02547 CL00113 |
| Прочие данные | |
| РНК тип | Ген ; rRNA |
| Domain(s) | Eukaryota ; |
| GO | 0005840 0003735 |
| SO | 0000652 |
| PDB структуры | PDBe |
Механизмы трансляции митохондрий и пластид (органеллы эндосимбиотического бактериального происхождения), и их бактериальные родственники имеют много общих черт, но также имеют заметные различия. Геномы органелл кодируют без исключения SSU и LSU рРНК, однако распределение генов 5S рРНК (rrn5) наиболее неравномерно. Rrn5 легко идентифицируется и встречается в геномах большинства пластид. Напротив, митохондриальный rrn5 изначально, по-видимому, ограничен растениями и небольшим количеством протистов. Дополнительные, более дивергентные органеллярные 5S рРНК были идентифицированы только с помощью специализированных ковариационных моделей, которые включают информацию о выраженном смещении состава последовательности и структурной вариации. Этот анализ выявил дополнительные гены 5S рРНК не только в митохондриальных геномах большинства протистских ветвей, но также и в геномах некоторых апикопластов (нефотосинтетических пластид патогенных простейших, таких как Toxoplasma gondii и Eimeria tenella ).
 Рисунок 4: Сравнение обычных и пермутированных моделей вторичной структуры 5S рРНК.
Рисунок 4: Сравнение обычных и пермутированных моделей вторичной структуры 5S рРНК. Митохондриальные 5S рРНК большинства страменопилов составляют наибольшее разнообразие вторичных структур. Пермутированные митохондриальные 5S рРНК в бурых водорослях представляют собой наиболее нетрадиционный случай, когда замыкающая спираль I, которая в противном случае объединяет 5 'и 3' концы молекулы, заменяется (закрытой) шпилькой, что приводит к открытый трехсторонний переход.
Текущие данные показывают, что митохондриальная ДНК только нескольких групп, в частности животных, грибов, альвеолят и эвгленозойных ген отсутствует. центральный выступ, в остальном занятый 5S рРНК и связанными с ней белками (см. рис. 2 ), был реконструирован различными способами. В митохондриальных рибосомах грибов 5S рРНК заменена последовательностями экспансии рРНК LSU. У кинетопластид (euglenozoans) центральный выступ полностью состоит из эволюционно новых митохондриальных рибосомных белков. Наконец, митохондриальные рибосомы животных кооперировали специфическую митохондриальную тРНК для замены отсутствующей 5S рРНК.