
Войти

Вторичная структура нуклеиновой кислоты представляет собой спаривание оснований в одном полимере нуклеиновой кислоты или между двумя полимерами. Его можно представить в виде списка оснований, которые спарены в молекуле нуклеиновой кислоты. Вторичные структуры биологических ДНК и РНК, как правило, различны: биологическая ДНК в основном существует в виде полностью спаренных двойных спиралей, в то время как биологическая РНК является одноцепочечной и часто образует сложные и сложные взаимодействия спаривания оснований из-за ее повышенной способности к образуют водородные связи, происходящие от дополнительной гидроксильной группы в сахаре рибозы.
В небиологическом контексте вторичная структура является жизненно важным фактором при проектировании нуклеиновых кислот структур нуклеиновых кислот для ДНК-нанотехнологий и ДНК-вычислений, поскольку характер спаривания оснований в конечном итоге определяет общую структуру молекул.

 Вверху: пара оснований AT, демонстрирующая две межмолекулярные водородные связи; внизу, пара оснований GC, демонстрирующая три межмолекулярные водородные связи. Основная статья: Базовая пара
Вверху: пара оснований AT, демонстрирующая две межмолекулярные водородные связи; внизу, пара оснований GC, демонстрирующая три межмолекулярные водородные связи. Основная статья: Базовая пара В молекулярной биологии два нуклеотида на противоположных комплементарных цепях ДНК или РНК, которые связаны водородными связями, называются парой оснований (часто сокращенно bp). В каноническом спаривании оснований Уотсона-Крика аденин (A) образует пару оснований с тимином (T), а гуанин (G) образует единицу с цитозином (C) в ДНК. В РНК тимин заменен урацилом (U). Альтернативные паттерны водородных связей, такие как колеблющаяся пара оснований и пара оснований Хугстина, также встречаются - особенно в РНК - давая начало сложным и функциональным третичным структурам. Важно отметить, что спаривание - это механизм, с помощью которого кодоны на молекулах информационной РНК распознаются антикодонами на транспортной РНК во время трансляции белка. Некоторые ДНК- или РНК-связывающие ферменты могут распознавать специфические паттерны спаривания оснований, которые идентифицируют определенные регуляторные области генов. Водородная связь - это химический механизм, лежащий в основе правил образования пар оснований, описанных выше. Соответствующее геометрическое соответствие доноров и акцепторов водородных связей позволяет стабильно образовываться только «правильным» парам. ДНК с высоким содержанием GC более стабильна, чем ДНК с низким содержанием GC, но вопреки распространенному мнению, водородные связи не стабилизируют ДНК в значительной степени, и стабилизация происходит в основном за счет взаимодействий стэкинга.
Более крупные азотистые основания, аденин и гуанин, являются членами класса химических структур с двойным кольцом, называемых пуринами ; более мелкие азотистые основания, цитозин и тимин (и урацил), являются членами класса однокольцевых химических структур, называемых пиримидинами. Пурины комплементарны только пиримидинам: пары пиримидин-пиримидин энергетически невыгодны, потому что молекулы находятся слишком далеко друг от друга для установления водородной связи; Пурин-пуриновые пары энергетически невыгодны, потому что молекулы расположены слишком близко, что приводит к перекрыванию отталкивания. Единственные другие возможные пары - GT и AC; эти спаривания не соответствуют друг другу, потому что структура доноров и акцепторов водорода не соответствует. Пара оснований колебания GU с двумя водородными связями действительно довольно часто встречается в РНК.
Гибридизация - это процесс связывания комплементарных пар оснований с образованием двойной спирали. Плавление - это процесс, при котором взаимодействие между цепями двойной спирали нарушается, разделяя две цепи нуклеиновой кислоты. Эти связи слабые, их легко разделить при легком нагревании, ферментах или физической силе. Плавление происходит предпочтительно в определенных точках нуклеиновой кислоты. Последовательности, богатые Т и А, легче расплавляются, чем богатые С и G. Конкретные основные стадии также чувствительны к плавлению ДНК, особенно стадии оснований TA и TG. Эти механические свойства отражаются в использовании последовательностей, таких как TATAA, в начале многих генов, чтобы помочь РНК-полимеразе плавить ДНК для транскрипции.
Разделение цепей мягким нагреванием, используемое в ПЦР, является простым при условии, что молекулы имеют менее примерно 10 000 пар оснований (10 пар оснований, или 10 т.п.н.). Переплетение нитей ДНК затрудняет разделение длинных сегментов. Клетка избегает этой проблемы, позволяя своим ДНК-плавящим ферментам ( геликазам ) работать одновременно с топоизомеразами, которые могут химически расщеплять фосфатный остов одной из цепей, так что она может вращаться вокруг другой. Геликазы раскручивают цепи, чтобы облегчить продвижение ферментов, считывающих последовательность, таких как ДНК-полимераза.
 Основные спиральные структуры нуклеиновых кислот (A-, B- и Z-форма)
Основные спиральные структуры нуклеиновых кислот (A-, B- и Z-форма) Вторичная структура нуклеиновой кислоты обычно делится на спирали (смежные пары оснований) и различные виды петель (неспаренные нуклеотиды, окруженные спиралями). Часто эти элементы или их комбинации, которые далее подразделяются на дополнительные категории, включая, например, tetraloops, псевдоузлы и стебель-петли.
Двойная спираль является важной третичной структурой в молекулах нуклеиновых кислот, которая тесно связана с вторичной структурой молекулы. Двойная спираль образована участками из множества следующих друг за другом пар оснований.
Двойная спираль нуклеиновой кислоты представляет собой спиральный полимер, обычно правый, содержащий две нуклеотидные цепи, которые соединяются в пары оснований. Один виток спирали составляет около десяти нуклеотидов и содержит большую бороздку и малую бороздку, причем большая бороздка шире, чем малая бороздка. Учитывая разницу в ширине большой бороздки и малой бороздки, многие белки, которые связываются с ДНК, делают это через более широкую большую бороздку. Возможны многие формы двойной спирали; для ДНК тремя биологически релевантными формами являются A-ДНК, B-ДНК и Z-ДНК, в то время как двойные спирали РНК имеют структуру, аналогичную A-форме ДНК.
 Вторичная структура стебель-петля РНК Основная статья: Стебель-петля
Вторичная структура стебель-петля РНК Основная статья: Стебель-петля Вторичная структура молекул нуклеиновой кислоты часто может быть однозначно разложена на стержни и петли. Структура стебель-петля (также часто называемая «шпилькой»), в которой спираль с парными основаниями заканчивается короткой непарной петлей, чрезвычайно распространена и является строительным блоком для более крупных структурных мотивов, таких как структуры клеверного листа, которые являются четырехспиральные соединения, подобные тем, которые встречаются в транспортной РНК. Также часто встречаются внутренние петли (короткая серия непарных оснований в более длинной спаренной спирали) и выпуклости (области, в которых одна цепь спирали имеет «лишние» вставленные основания без аналогов в противоположной цепи).
Есть много элементов вторичной структуры, имеющих функциональное значение для биологических РНК; некоторыми известными примерами являются Rho-независимые терминаторные петли-ножки и тРНК клеверный лист. Продолжаются активные исследования по определению вторичной структуры молекул РНК с подходами, включающими как экспериментальные, так и вычислительные методы (см. Также Список программ для предсказания структуры РНК ).
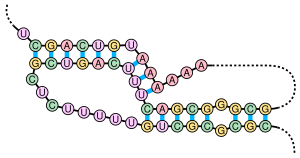 Структура псевдоузла РНК. Например, РНК-компонент теломеразы человека.
Структура псевдоузла РНК. Например, РНК-компонент теломеразы человека. Псевдоузел представляет собой вторичную структуру нуклеиновой кислоты, содержащую, по меньшей мере, две структуры « стебель-петля», в которых половина одного стебля вставлена между двумя половинами другого стебля. Псевдоузлы складываются в узловые трехмерные конформации, но не являются настоящими топологическими узлами. Спаривание оснований в псевдоузлах не очень хорошо вложенные; то есть встречаются пары оснований, которые «перекрывают» друг друга в положении последовательности. Это делает невозможным предсказать присутствие общих псевдоузлов в последовательностях нуклеиновых кислот стандартным методом динамического программирования, который использует рекурсивную систему подсчета очков для идентификации парных основ и, следовательно, не может обнаруживать невложенные пары оснований с помощью общих алгоритмов. Однако ограниченные подклассы псевдоузлов можно предсказать с помощью модифицированных динамических программ. Новые методы предсказания структуры, такие как стохастические контекстно-свободные грамматики, также не могут учитывать псевдоузлы.
Псевдоузлы могут образовывать множество структур с каталитической активностью, а некоторые важные биологические процессы зависят от молекул РНК, которые образуют псевдоязычные узлы. Например, РНК-компонент теломеразы человека содержит псевдоузел, который имеет решающее значение для его активности. Рибозим вируса гепатита дельта является хорошо известным примером каталитической РНК с псевдоузлом в ее активном центре. Хотя ДНК также может образовывать псевдоузлы, они обычно не присутствуют в стандартных физиологических условиях.
Большинство методов предсказания вторичной структуры нуклеиновых кислот основаны на термодинамической модели ближайшего соседа. Обычный метод определения наиболее вероятных структур с учетом последовательности нуклеотидов использует алгоритм динамического программирования, который стремится найти структуры с низкой свободной энергией. Алгоритмы динамического программирования часто запрещают псевдоузлы или другие случаи, когда пары оснований не полностью вложены, поскольку рассмотрение этих структур становится очень дорогостоящим в вычислительном отношении даже для небольших молекул нуклеиновых кислот. Другие методы, такие как стохастические контекстно-свободные грамматики, также могут быть использованы для предсказания вторичной структуры нуклеиновой кислоты.
Для многих молекул РНК вторичная структура очень важна для правильного функционирования РНК - часто даже больше, чем фактическая последовательность. Этот факт помогает при анализе некодирующих РНК, иногда называемых «генами РНК». Одно приложение биоинформатики использует предсказанные вторичные структуры РНК для поиска в геноме некодирующих, но функциональных форм РНК. Например, микроРНК имеют канонические длинные структуры «стебель-петля», прерванные небольшими внутренними петлями.
Вторичная структура РНК применяется при сплайсинге РНК у некоторых видов. У людей и других четвероногих было показано, что без белка U2AF2 процесс сплайсинга подавляется. Однако, в данио и других костистых сплайсинг РНК процесс все еще может иметь место на определенных генов в отсутствие U2AF2. Это может быть связано с тем, что 10% генов у рыбок данио имеют чередующиеся пары оснований TG и AC в 3'-сайте сплайсинга (3'ss) и 5'-сайте сплайсинга (5'ss) соответственно на каждом интроне, что изменяет вторичную структуру РНК. Это говорит о том, что вторичная структура РНК может влиять на сплайсинг, возможно, без использования белков, подобных U2AF2, которые, как считалось, необходимы для сплайсинга.
Вторичная структура РНК может быть определена по координатам атомов (третичная структура), полученным с помощью рентгеновской кристаллографии, которые часто хранятся в банке данных о белках. Текущие методы включают 3DNA / DSSR и MC-annotate.