
Войти
| WRKY | |||||||||
|---|---|---|---|---|---|---|---|---|---|
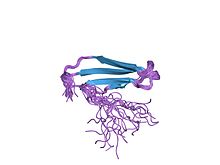 структура решения c-концевого домена wrky atwrky4 структура решения c-концевого домена wrky atwrky4 | |||||||||
| Идентификаторы | |||||||||
| Символ | WRKY | ||||||||
| Pfam | PF03106 | ||||||||
| Pfam клан | CL0274 | ||||||||
| InterPro | IPR003657 | ||||||||
| |||||||||
Домен WRKY находится в семействе факторов транскрипции WRKY, классе факторов транскрипции. Домен WRKY обнаружен почти исключительно в растениях, хотя гены WRKY, по-видимому, присутствуют у некоторых дипломонад, социальных амеб и других амебозоидов и грибов incertae sedis. Они отсутствуют у других нерастительных видов. Факторы транскрипции WRKY были важной областью исследований растений в течение последних 20 лет. ДНК-связывающий домен WRKY распознает W-бокс (T) TGAC (C / T) (и варианты этой последовательности) цис-регуляторный элемент.
Факторы транскрипции WRKY содержат один или два домена белка WRKY. Белковый домен WRKY представляет собой ДНК-связывающий домен длиной от 60 до 70 аминокислот. Домен характеризуется высококонсервативным основным мотивом WRKYGQK и областью цинкового пальца. Домены цинковых пальцев цистеин и гистидин имеют тип CX 4-5 CX 22-23 HXH или CX 7CX23HXC., где X может быть любой аминокислотой. Цинковый палец связывает ион Zn, который необходим для функционирования белка. Хотя WRKYGQK является высококонсервативным в большинстве доменов WRKY, изменения в основной последовательности были задокументированы. Часто встречающимся вариантом основной последовательности является WRKYGKK, который присутствует у большинства видов растений.
Структура домена белка WRKY была впервые определена в 2005 году с использованием ядерного магнитного резонанса (ЯМР) а позже кристаллография. Белковый домен WRKY имеет глобулярную форму, состоящую из пяти антипараллельных β-тяжей. Основной мотив WRKYGQK находится на второй β-цепи. Восемнадцать аминокислот являются высококонсервативными в домене белка WRKY, включая основной мотив, цистеины и гистидины, связывающиеся с цинковыми пальцами, и триаду, образующую солевой мостик DWK. Триада состоит из консервативного триптофана (W) основного мотива, наряду с аспарагиновой кислотой (D) на четыре аминокислоты выше и лизин (K) на 29 аминокислот ниже по течению. это, стабилизируя весь домен. Пять аминокислот на третьей β-цепи (PRSYY) также хорошо консервативны в домене WRKY. Важно отметить, что гены WRKY содержат консервативный интрон в домене WRKY, который находится в месте, кодирующем PR аминокислотной последовательности PRSYY, что объясняет консервативность этого мотива.
Домен WRKY образует уникальную клиновидную структуру, которая входит перпендикулярно в большую бороздку цепи ДНК. Домены белка WRKY взаимодействуют с цис-элементом (T / A) TGAC (T / A), также называемым W-боксом. Последние данные свидетельствуют о том, что ядро GAC W-бокса является основной мишенью домена WRKY, а фланкирующие последовательности помогают определять взаимодействие ДНК с очень специфическими белками WRKY. Остатки RKYGQK основного мотива и дополнительные аргинин и лизиновые остатки домена WRKY ответственны за взаимодействие с фосфатным остовом семи последовательных пар оснований ДНК, включая ядро GAC. Замена триптофана, тирозина или любого лизина мотива WRKYGQK на аланин полностью устраняет связывание ДНК, указывая на то, что эти аминокислоты необходимы для распознавания элемента W-бокса. Хотя это не является существенным, изменение мотива WRKYGQK аргинина, глицина или глутамина на аланин снижает связывание ДНК с W-боксом. В целом, эти сложные взаимодействия белкового домена WRKY с ДНК приводят к активации генов, необходимой для многих аспектов развития и защиты растений.