
Войти
| Просмотр / редактирование Человека | Просмотр / редактирование мыши |
Сигнальный белок агути - это белок, который у человека кодируется геном ASIP . Он отвечает за распределение пигмента меланин у млекопитающих. Агути взаимодействует с рецептором меланокортина, чтобы определить, производит ли меланоцит (пигментная клетка) феомеланин (пигмент от красного до желтого) или эумеланин (пигмент от коричневого до черного). Это взаимодействие отвечает за образование четких светлых и темных полос в волосах животных, таких как агути, в честь которого назван ген. У других видов, таких как лошади, передача сигналов агути отвечает за определение того, какие части тела будут красными или черными. Мыши с агути дикого типа будут серыми, причем каждый волос будет частично желтым, а частично черным. Мутации потери функции у мышей и других видов вызывают черную окраску шерсти, в то время как мутации, вызывающие экспрессию во всем теле у мышей, вызывают желтый мех и ожирение.
Сигнальный белок агути (ASIP) является конкурентным антагонист с альфа-меланоцитостимулирующим гормоном (α-MSH) для связывания с рецептором меланокортина 1 (MC1R) белками. Активация α-MSH вызывает выработку более темного эумеланина, в то время как активация ASIP вызывает выработку более красного феомеланина. Это означает, что там, где и пока выражается агути, растущая часть волос будет желтой, а не черной.
У мышей ген агути кодирует паракрин сигнальную молекулу, которая заставляет волосяной фолликул меланоциты синтезировать желтый пигмент феомеланин вместо черного или коричневого пигмента эумеланина. Плейотропные эффекты конститутивной экспрессии мышиного гена включают ожирение у взрослых, повышенную восприимчивость опухоли и преждевременное бесплодие. Этот ген очень похож на ген мыши и кодирует секретируемый белок, который может (1) влиять на качество волос пигментацию, (2) действовать как обратный агонист гормона, стимулирующего альфа-меланоциты., (3) играют роль в нейроэндокринных аспектах действия меланокортина и (4) играют функциональную роль в регулировании липидного обмена в адипоцитах.
в У мышей аллель дикого типа агути (A) представляет собой серый фенотип, однако многие варианты аллелей были идентифицированы с помощью генетического анализа, что привело к широкому диапазону фенотипов, отличных от типичной серой шерсти. Наиболее широко изученными вариантами аллелей являются летальная мутация желтого цвета (А) и жизнеспособная мутация желтого цвета (А), которые вызваны эктопической экспрессией агути. Эти мутации также связаны с синдромом желтого ожирения, который характеризуется ранним началом ожирением, гиперинсулинемией и туморогенезом. Локус гена мыши агути обнаружен на хромосоме 2 и кодирует белок из 131 аминокислоты. Этот белок сигнализирует о распределении пигментов меланина в эпителиальных меланоцитах, расположенных в основании волосяных фолликулов, причем экспрессия более чувствительна на вентральных волосах, чем на спинных. Агути не секретируется непосредственно в меланоцитах, поскольку он действует как паракринный фактор на клетки сосочков дермы, подавляя высвобождение меланокортина. Меланокортин действует на фолликулярные меланоциты, увеличивая выработку эумеланина, пигмента меланина, отвечающего за каштановые и черные волосы. При экспрессии агути преобладает выработка феомеланина, пигмента меланина, который дает желтый или красный цвет волос.
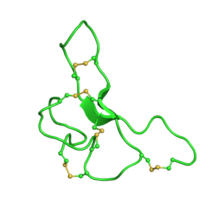 Семейство структур ЯМР сигнального белка агути, С-концевой узел узла. Запись PDB 1y7k
Семейство структур ЯМР сигнального белка агути, С-концевой узел узла. Запись PDB 1y7k сигнальный пептид Агути принимает мотив цистинового узла ингибитора. Наряду с гомологичным родственным агути пептидом, это единственные известные белки млекопитающих, которые принимают эту складку. Пептид состоит из 131 аминокислоты.
Смертельная мутация желтого цвета (А) была первой эмбриональной мутацией, охарактеризованной у мышей, поскольку гомозиготные летальные мыши желтого цвета (А / А) умирают на ранней стадии развития из-за ошибки в дифференцировке трофэктодермы. Смертельные желтые гомозиготы сегодня редки, в то время как летальные желтые и жизнеспособные желтые гетерозиготы (A / a и A / a) остаются более распространенными. У мышей дикого типа агути экспрессируется в коже только во время роста волос, но эти доминантные мутации желтого цвета вызывают его экспрессию и в других тканях. Эта эктопическая экспрессия гена агути связана с синдромом желтого ожирения, который характеризуется ранним началом ожирением, гиперинсулинемией и туморогенезом.
Смертельным Желтая (A) мутация возникает из-за вышестоящей делеции в стартовом сайте транскрипции агути. Эта делеция приводит к потере геномной последовательности агути, за исключением промотора и первого некодирующего экзона Raly, широко экспрессируемого гена у млекопитающих. Кодирующие экзоны агути находятся под контролем промотора Рали, инициируя повсеместную экспрессию агути, увеличивая продукцию феомеланина по сравнению с эумеланином и приводя к развитию желтого фенотипа.
 Предлагаемый механизм взаимосвязи между эктопической экспрессией агути и развитием синдрома желтого ожирения
Предлагаемый механизм взаимосвязи между эктопической экспрессией агути и развитием синдрома желтого ожирения Жизнеспособная мутация желтого цвета (A) обусловлена изменением длины мРНК агути, так как экспрессируемый ген становится длиннее нормальной длины гена агути. Это вызвано вставкой ретротранспозона внутрицистернальной А-частицы (IAP) перед стартовым сайтом транскрипции агути. Затем на проксимальном конце гена неизвестный промотор вызывает конституциональную активацию агути и появление у индивидов фенотипа, соответствующего летальной мутации желтого цвета. Хотя механизм активации промотора, контролирующего жизнеспособную мутацию желтого цвета, неизвестен, интенсивность окраски шерсти коррелирует со степенью метилирования гена , которая определяется рационом матери и воздействием окружающей среды. Поскольку агути сам по себе подавляет рецепторы меланокортина, ответственные за выработку эумеланина, фенотип желтого цвета усугубляется как при летальной, так и при жизнеспособной желтой мутации по мере увеличения экспрессии гена агути. Агути уникален, потому что, хотя это рецессивный аллель, гетерозиготы будут выглядеть желтыми, а не доминирующими коричневыми или черными.
Жизнеспособные гетерозиготы желтого (A / a) и смертоносного (A / a) желтого (A / a) имеют более короткую продолжительность жизни и повышенный риск развития раннего ожирения, сахарного диабета 2 типа и различных опухолей. Повышенный риск развития ожирения связан с нарушением регуляции аппетита, поскольку агути агонизирует связанный с агути белок (AGRP), ответственный за стимуляцию аппетита через NPY / AGRP или орексигенные нейроны гипоталамуса. Агути также способствует ожирению за счет антагонизма меланоцит-стимулирующего гормона (MSH) на рецепторе меланокортина (MC4R), поскольку MC4R отвечает за регулирование приема пищи путем подавления сигналов аппетита. Повышение аппетита связано с изменениями в метаболизме питательных веществ из-за паракринного действия агути на жировую ткань, увеличения уровней печеночного липогенеза, снижения уровней липолиза и увеличение гипертрофии адипоцитов. Это увеличивает массу тела и приводит к трудностям с потерей веса, поскольку метаболические пути нарушаются. Гиперинсулинемия вызвана мутациями агути, поскольку белок агути функционирует кальций-зависимым образом, увеличивая секрецию инсулина в бета-клетках поджелудочной железы, увеличивая риски резистентности к инсулину. Повышенное образование опухолей происходит из-за повышенной митотической скорости агути, которые локализуются в эпителиальных и мезенхимальных тканях.
 Эти мыши генетически идентичны, несмотря на то, что выглядят фенотипически разными. Мыши слева кормили бисфенолом А (BPA) с нормальной диетой для мышей, а мышь справа кормили BPA с пищей, богатой метилом. Левая мышь желтая и тучная, а правая коричневая и здоровая.
Эти мыши генетически идентичны, несмотря на то, что выглядят фенотипически разными. Мыши слева кормили бисфенолом А (BPA) с нормальной диетой для мышей, а мышь справа кормили BPA с пищей, богатой метилом. Левая мышь желтая и тучная, а правая коричневая и здоровая. Для правильного функционирования агути необходимо метилирование ДНК. Метилирование происходит в шести богатых гуанин-цитозином (GC) последовательностях в 5 ’длинном концевом повторе элемента IAP в жизнеспособной желтой мутации. Метилирование гена приводит к тому, что ген не экспрессируется, потому что это приводит к отключению промотора . В утробе матери диета матери может вызвать метилирование или деметилирование. Когда эта область неметилирована, происходит эктопическая экспрессия агути, и проявляются желтые фенотипы, потому что феомеланин экспрессируется вместо эумеланина. Когда область метилирована, агути экспрессируется нормально, и встречаются серый и коричневый фенотипы (эумеланин). Эпигенетическое состояние элемента IAP определяется уровнем метилирования, поскольку индивидуумы демонстрируют широкий спектр фенотипов в зависимости от степени метилирования ДНК. Повышенное метилирование коррелирует с повышенной экспрессией нормального гена агути. Низкие уровни метилирования могут вызывать импринтинг гена, что приводит к тому, что потомство демонстрирует последовательные фенотипы своих родителей, поскольку эктопическая экспрессия агути наследуется через негеномные механизмы.
Метилирование ДНК определяется в утробе матери. питанием матери и воздействием окружающей среды. Метил синтезируется de novo, но достигается через диету с помощью фолиевой кислоты, метионина, бетаина и холина, поскольку эти питательные вещества участвуют в последовательном метаболическом пути синтеза метила. Достаточное количество цинка и витамина B12 необходимы для синтеза метила, поскольку они действуют как кофакторы для переноса метильных групп.
Когда недостаточное количество метила доступно во время раннего эмбрионального развития, метилирование ДНК не может произойти, что увеличивает эктопическую экспрессию агути и приводит к представлению фенотипов летального желтого и жизнеспособного желтого, которые сохраняются во взрослой жизни. Это приводит к развитию синдрома желтого ожирения, который нарушает нормальное развитие и увеличивает склонность к развитию хронических заболеваний. Обеспечение материнского рациона высоким содержанием метиловых эквивалентов является ключевой профилактической мерой для снижения эктопической экспрессии агути у потомства. Вмешательство в диету с помощью добавления метила снижает импринтинг в локусе агути, поскольку повышенное потребление метила вызывает полное метилирование элемента IAP и снижение эктопической экспрессии агути. Это снижает долю потомства с желтым фенотипом и увеличивает количество потомков, которые напоминают мышей дикого типа агути с серой шерстью. Две генетически идентичные мыши могли выглядеть очень по-разному фенотипически из-за рациона матери, когда мыши находились в утробе матери. Если у мышей есть ген агути, он может быть выражен из-за того, что мать придерживается типичной диеты, и у потомства будет желтая шерсть. Если бы та же мать ела диету, богатую метилом, с добавлением цинка, витамина B12 и фолиевой кислоты, тогда ген агути у потомства, вероятно, стал бы метилированным, он не проявился бы, и вместо этого цвет шерсти был бы коричневым. У мышей желтый цвет шерсти также связан с проблемами со здоровьем у мышей, включая ожирение и диабет.
сигнальный белок агути (ASP) является человеческим гомологом мышиного агути. Он кодируется геном агути человека на хромосоме 20 и представляет собой белок, состоящий из 132 аминокислот. Он экспрессируется гораздо шире, чем мышиный агути, и обнаруживается в жировой ткани, поджелудочной железе, семенниках и яичниках, тогда как мышиный агути экспрессируется исключительно в меланоцитах. ASP имеет 85% сходство с мышиной формой агути. Поскольку эктопическая экспрессия мышиного агути приводит к развитию синдрома желтого ожирения, ожидается, что это будет устойчиво у людей. Синдром желтого ожирения увеличивает развитие многих хронических заболеваний, включая ожирение, сахарный диабет II типа и онкогенез.
ASP имеет фармакологическую активацию, аналогичную мышиной агути, поскольку рецепторы меланокортина ингибируются посредством конкурентного антагонизма. Ингибирование меланокортина с помощью ASP также может осуществляться неконкурентными методами, что расширяет спектр его эффектов. Функция ASP отличается от мышиного агути. ASP влияет на качество пигментации волос, тогда как мышиный агути контролирует распределение пигментов, определяющих цвет шерсти. ASP выполняет нейроэндокринные функции, соответствующие мышиному агути, поскольку он агонизирует через нейроны AgRP в гипоталамусе и противодействует MSH в MC4R, которые снижают сигналы сытости. AgRP действует как стимулятор аппетита и увеличивает аппетит, снижая метаболизм. Из-за этих механизмов AgRP может быть связан с увеличением массы тела и ожирением как у людей, так и у мышей. Сверхэкспрессия AgRP была связана с ожирением у мужчин, в то время как определенные полиморфизмы AgRP были связаны с расстройствами пищевого поведения, такими как нервная анорексия. Механизм, лежащий в основе гиперинсулинемии у людей, согласуется с мышиным агути, поскольку секреция инсулина повышается за счет чувствительной к кальцию передачи сигналов в бета-клетках поджелудочной железы. Механизм индуцированного ASP туморогенеза у людей остается неизвестным.
Эта статья включает текст из Национальной медицинской библиотеки США, которая находится в общественном достоянии.

