
Войти
 Ксилема (синяя) транспортирует воду и минералы от корней вверх.
Ксилема (синяя) транспортирует воду и минералы от корней вверх. Ксилема является одним из двух типов транспорта ткани в сосудистых растениях, а флоэма является другим. Основная функция ксилемы - транспортировать воду от корней к стеблям и листьям, но она также транспортирует питательные вещества. Слово «ксилема» происходит от греческого слова ξύλον (ксилон), что означает «дерево»; Самая известная ткань ксилемы - это древесина, хотя она встречается повсюду в растении. Термин был введен Карлом Нэгели в 1858 году.
 Поперечное сечение некоторых ячеек ксилемы
Поперечное сечение некоторых ячеек ксилемы Наиболее характерные ксилемы ячейки представляют собой длинные трахеальные элементы, переносящие воду. трахеиды и сосудистые элементы различаются по своей форме; элементы сосудов короче и соединены в длинные трубки, которые называются сосудами.
Ксилема также содержит два других типа клеток: паренхима и волокна.
Можно найти ксилему :
на переходных стадиях развития растений с вторичным ростом первые две категории не исключают друг друга, хотя обычно сосудистый пучок будет содержать только первичную ксилему.
Паттерн ветвления, демонстрируемый ксилемой, следует закону Мюррея.
Первичная ксилема образуется во время первичного роста из прокамбия. Он включает протоксилему и метаксилему. Метаксилема развивается после протоксилемы, но до вторичной ксилемы. Метаксилема имеет более широкие сосуды и трахеиды, чем протоксилема.
Вторичная ксилема образуется во время вторичного роста из сосудистого камбия. Хотя вторичная ксилема также обнаруживается у представителей групп голосеменных Gnetophyta и Ginkgophyta и в меньшей степени у представителей Cycadophyta, две основные группы, в которых может быть обнаружена вторичная ксилема:
Ксилема, сосуды и трахеиды корней, стеблей и т. Д. листья соединены между собой, образуя непрерывную систему водопроводящих каналов, идущих ко всем частям растений. Система транспортирует воду и растворимые минеральные питательные вещества от корней по всему растению. Он также используется для восполнения потери воды во время транспирации и фотосинтеза. Ксилема сок состоит в основном из воды и неорганических ионов, хотя он также может содержать ряд органических химикатов. Транспорт является пассивным, он не питается энергией, расходуемой самими элементами трахеи, которые умерли от зрелости и больше не имеют живого содержимого. Транспортировка сока вверх становится более трудной, поскольку высота растения увеличивается, и считается, что транспортировка воды ксилемой вверх ограничивает максимальную высоту деревьев. Три явления вызывают течь ксилемного сока:
Первичной силой, которая создает капиллярное действие движение воды вверх у растений, является адгезия между водой и поверхностью каналов ксилемы. Капиллярное действие обеспечивает силу, которая устанавливает равновесную конфигурацию, уравновешивая силу тяжести. Когда транспирация удаляет воду из верхней части, поток необходим, чтобы вернуться к равновесию.
Транспирационное притяжение возникает в результате испарения воды с поверхностей ячеек в листьях. Это испарение заставляет поверхность воды проникать в поры в клеточной стенке. За счет капиллярного действия вода образует вогнутые мениски внутри пор. Высокое поверхностное натяжение воды вытягивает вогнутость наружу, создавая достаточную силу, чтобы поднимать воду на высоту ста метров от уровня земли до самого высокого дерева . ветви.
Транспирационная тяга требует, чтобы сосуды, транспортирующие воду, были очень маленькими в диаметре; в противном случае кавитация нарушит толщину воды. И по мере того, как вода испаряется из листьев, растение вытягивает больше воды, чтобы заменить ее. Когда давление воды внутри ксилемы достигает предельного уровня из-за низкого поступления воды из корней (например, если почва сухая), газы выходят из раствора и образуют пузырь - эмболия формы, которые будут быстро распространяться на другие соседние клетки, если не присутствуют окаймленные ямки (они имеют заглушку, называемую торцом, которая закрывает отверстие между соседними клетками и останавливает распространение эмболии). Даже после того, как произошла эмболия, растения способны восполнить ксилему и восстановить функциональность.
Теория сцепления-напряжения - это теория межмолекулярное притяжение, которое объясняет процесс потока воды вверх (против силы силы тяжести ) через ксилему растений. Он был предложен в 1894 году Джоном Джоли и Генри Горацио Диксоном. Несмотря на многочисленные возражения, это наиболее широко распространенная теория переноса воды через сосудистую систему растений, основанная на классических исследованиях Диксона-Джоли (1894 г.), Юджена Аскенази (1845–1903 гг.) (1895 г.) и Диксона (1914 г. 1924).
Вода - это полярная молекула. Когда две молекулы воды приближаются друг к другу, слегка отрицательно заряженный атом кислорода в одной образует водородную связь со слегка положительно заряженным атомом водорода в другом. Эта сила притяжения, наряду с другими межмолекулярными силами, является одним из основных факторов, ответственных за возникновение поверхностного натяжения в жидкой воде. Это также позволяет растениям вытягивать воду от корня через ксилему к листу.
Вода постоянно теряется из-за испарения листьев. Когда одна молекула воды теряется, другую затягивают процессы сцепления и напряжения. Тяга при транспирации, использующая капиллярное действие и естественное поверхностное натяжение воды, является основным механизмом движения воды в растениях. Однако это не единственный задействованный механизм. Любое использование воды в листьях заставляет воду перемещаться в них.
Транспирация в листьях создает напряжение (перепад давления) в клеточных стенках клеток мезофилла. Из-за этого напряжения вода поднимается от корней к листьям, чему способствует когезия (притяжение между отдельными молекулами воды из-за водородных связей ) и адгезия. (липкость между молекулами воды и гидрофильными клеточными стенками растений). Этот механизм потока воды работает из-за потенциала воды (вода течет от высокого к низкому потенциалу) и правил простой диффузии.
За последнее столетие было много исследование механизма транспорта ксилемного сока; сегодня большинство ученых-растениеводов продолжают соглашаться с тем, что теория сцепления-напряжения лучше всего объясняет этот процесс, но были предложены теории множества сил, которые предполагают несколько альтернативных механизмов, включая продольные клеточные и ксилемные градиенты осмотического давления >, осевые градиенты потенциала в сосудах, а не гель-и газо-пузырьковый поддерживают межфазных градиенты.
 диаграмма, показывающая настройку в бомбы давления
диаграмма, показывающая настройку в бомбы давления До недавнего времени, дифференциальное давление (всасывание) из transpirational тяги может быть измерено лишь косвенно, путем применения внешнего давления с бомба давления, чтобы противодействовать этому. Когда была разработана технология выполнения прямых измерений с помощью датчика давления, первоначально возникли некоторые сомнения в правильности классической теории, поскольку некоторые исследователи не смогли продемонстрировать отрицательное давление. Более поздние измерения, по большей части, подтверждают классическую теорию. Перенос ксилемы управляется комбинацией транспирационного притяжения сверху и корневого давления снизу, что усложняет интерпретацию измерений.
Ксилема возникла в начале истории наземной растительной жизни. Ископаемые растения с анатомически сохранившейся ксилемой известны с силурия (более 400 миллионов лет назад), а следы окаменелостей, напоминающие отдельные клетки ксилемы, можно найти в более ранних породах ордовика. Самая ранняя настоящая и узнаваемая ксилема состоит из трахеид со спирально-кольцевым армирующим слоем, добавленным к клеточной стенке. Это единственный тип ксилемы, обнаруженный у самых ранних сосудистых растений, и этот тип клеток продолжает обнаруживаться в протоксилеме (первой сформированной ксилеме) всех живых групп сосудистых растений. Несколько групп растений позже независимо развили без ямок трахеидных клеток в результате конвергентной эволюции. У живых растений трахеиды с косточкой не появляются в развитии до созревания метаксилемы (следующей за протоксилемой).
У большинства растений без косточки трахеиды функционируют как первичные транспортные клетки. Другой тип сосудистого элемента, обнаруженный у покрытосеменных, - это сосудистый элемент. Элементы сосуда соединяются встык, образуя сосуды, в которых вода течет беспрепятственно, как в трубе. Наличие ксилемных сосудов считается одним из ключевых нововведений, которые привели к успеху покрытосеменных. Однако наличие сосудистых элементов не ограничивается покрытосеменными, и они отсутствуют в некоторых архаичных или «базальных» ветвях покрытосеменных: (например, Amborellaceae, Tetracentraceae, Trochodendraceae и Winteraceae ), а их вторичная ксилема описана Артуром Кронквистом как «примитивно бессосудистые». Кронквист считал, что сосуды Gnetum сходятся с сосудами покрытосеменных. Является ли отсутствие сосудов у базальных покрытосеменных примитивным условием, это оспаривается, альтернативная гипотеза утверждает, что элементы сосудов произошли от предшественников покрытосеменных и впоследствии были потеряны.
 Фотографии, показывающие элементы ксилемы в побеге фигового дерева (Ficus alba): измельченные в соляной кислоте между предметными стеклами и покровными стеклами
Фотографии, показывающие элементы ксилемы в побеге фигового дерева (Ficus alba): измельченные в соляной кислоте между предметными стеклами и покровными стеклами Для фотосинтеза растения должны поглощать CO. 2 из атмосферы. Однако за это приходится платить: в то время как устьицы открыты для проникновения CO. 2, вода может испаряться. Вода теряется намного быстрее, чем поглощается CO. 2, поэтому растениям необходимо ее восполнить, и они разработали системы для транспортировки воды из влажной почвы к месту фотосинтеза. Ранние растения засасывали воду между стенками своих клеток, а затем развили способность контролировать потерю воды (и накопление CO. 2) с помощью устьиц. Специализированные водные транспортные ткани вскоре превратились в гидроидов, трахеид, затем вторичную ксилему, затем энтодерму и, наконец, сосуды.
Высокие уровни CO. 2 в силурийско-девонские времена, когда впервые появились растения. колонизация земли означала, что потребность в воде была относительно низкой. По мере того как CO. 2 выводился из атмосферы растениями, при его захвате терялось больше воды, и развивались более элегантные механизмы переноса. По мере развития механизмов переноса воды и водонепроницаемой кутикулы растения могли выжить, не будучи постоянно покрытыми пленкой воды. Этот переход от пойкилогидрии к гомойогидрии открыл новый потенциал для колонизации. Затем растениям нужна была прочная внутренняя структура, которая содержала длинные узкие каналы для транспортировки воды из почвы ко всем различным частям надземного растения, особенно к тем частям, где происходил фотосинтез.
Во времена силурия CO. 2 был легко доступен, поэтому для его получения требовалось немного воды. К концу каменноугольного периода, когда уровни CO. 2 снизились до уровня, близкого к сегодняшнему, на единицу поглощения CO. 2 терялось примерно в 17 раз больше воды. Однако даже в эти "легкие" ранние дни вода была в дефиците, и ее приходилось транспортировать к частям растения из влажной почвы, чтобы избежать высыхания. В этом раннем водном транспорте использовался механизм сцепления-напряжения, присущий воде. Вода имеет тенденцию диффундировать в более сухие области, и этот процесс ускоряется, когда вода может быть направлена вдоль ткани с небольшими промежутками. В небольших проходах, например, между стенками растительных клеток (или в трахеидах), столб воды ведет себя как резина - когда молекулы испаряются с одного конца, они тянут молекулы за собой по каналам. Следовательно, только транспирация была движущей силой переноса воды у ранних растений. Однако без специальных транспортных сосудов механизм сцепления-натяжения не может транспортировать воду более чем примерно на 2 см, что сильно ограничивает размер самых ранних растений. Этот процесс требует постоянной подачи воды с одного конца для поддержания цепей; Чтобы не истощать его, растения создали водонепроницаемую кутикулу. Ранняя кутикула могла не иметь пор, но не покрывала всю поверхность растения, так что газообмен продолжался. Однако иногда обезвоживание было неизбежным; Ранние растения справляются с этим за счет того, что между их клеточными стенками хранится много воды, а когда дело доходит до этого, они выдерживают трудные времена, задерживая жизнь, пока не поступит больше воды.
 A трубка с полосами поздний силурий / ранний девон. Полосы на этом образце трудно увидеть, так как непрозрачное углеродистое покрытие скрывает большую часть трубки. Полосы видны только в некоторых местах в левой половине изображения - щелкните изображение, чтобы увеличить его. Масштабная линейка: 20 мкм
A трубка с полосами поздний силурий / ранний девон. Полосы на этом образце трудно увидеть, так как непрозрачное углеродистое покрытие скрывает большую часть трубки. Полосы видны только в некоторых местах в левой половине изображения - щелкните изображение, чтобы увеличить его. Масштабная линейка: 20 мкм Чтобы освободиться от ограничений небольшого размера и постоянной влажности, создаваемых паренхиматической транспортной системой, растениям требовалась более эффективная водная транспортная система. В течение раннего силурия они разработали специализированные клетки, которые были одревесневшими (или содержали аналогичные химические соединения), чтобы избежать имплозии; этот процесс совпал с гибелью клеток, позволяя опустошить их внутренности и пропустить через них воду. Эти более широкие, мертвые, пустые клетки были в миллион раз более проводящими, чем метод межклеточного взаимодействия, давая возможность для переноса на большие расстояния и более высокие скорости диффузии CO. 2.
Самые ранние макрофоссилии, несущие водопроводные трубы, - это силурийские растения, относящиеся к роду Cooksonia. Раннедевонские претрахеофиты Aglaophyton и Horneophyton имеют структуру, очень похожую на гидроиды современных мхов. Растения продолжали изобретать новые способы уменьшения сопротивления потоку внутри своих клеток, тем самым повышая эффективность водного транспорта. Полосы на стенках трубок, которые на самом деле появляются с раннего силурия и далее, являются ранней импровизацией, способствующей легкому течению воды. Полосчатые трубки, а также трубки с ямчатым орнаментом на стенках были одревесневшими и, когда они образуют одноклеточные каналы, считаются трахеидами. Эти конструкции транспортных ячеек «следующего поколения» имеют более жесткую конструкцию, чем гидроиды, что позволяет им справляться с более высокими уровнями давления воды. Трахеиды могут иметь единое эволюционное происхождение, возможно, внутри роголистников, объединяя все трахеофиты (но они могли эволюционировать более одного раза).
Водный транспорт требует регулирования, а динамический контроль обеспечивается устьицами. Регулируя количество газообмена, они могут ограничить количество воды, теряемой при транспирации. Это важная роль там, где водоснабжение не является постоянным, и действительно, кажется, что устьица развились до трахеид, присутствуя у несосудистых роголистников.
энтодерма, вероятно, возникла во время Силу. -Девон, но первые ископаемые свидетельства такой структуры относятся к каменноугольному периоду. Эта структура в корнях покрывает ткань, транспортирующую воду, и регулирует ионный обмен (и предотвращает попадание нежелательных патогенов и т. Д. В систему транспорта воды). Энтодерма также может оказывать давление вверх, вытесняя воду из корней, когда транспирации недостаточно.
Как только растения развили этот уровень контролируемого переноса воды, они стали действительно гомоатомными, способны извлекать воду из окружающей среды через корневидные органы, а не полагаться на пленку поверхностной влаги, что позволяет им расти до больший размер. В результате их независимости от окружающей среды они потеряли способность выживать при высыхании - дорогостоящая черта, которую нужно сохранить.
В течение девона максимальный диаметр ксилемы увеличивался со временем, а минимальный диаметр оставался довольно постоянным. К среднему девону диаметр трахеид некоторых линий растений (Zosterophyllophytes ) вышел на плато. Более широкие трахеиды позволяют воде транспортироваться быстрее, но общая скорость транспортировки зависит также от общей площади поперечного сечения самого пучка ксилемы. Увеличение толщины сосудистых пучков, по-видимому, также коррелирует с шириной осей растений и высотой растений; это также тесно связано с внешним видом листьев и повышенной плотностью устьиц, что увеличивает потребность в воде.
Хотя более широкие трахеиды с прочными стенками позволяют достичь более высокого давления переноса воды, это увеличивает проблема кавитации. Кавитация возникает, когда внутри сосуда образуется пузырь воздуха, который разрывает связи между цепочками молекул воды и не позволяет им поднимать больше воды за счет своего когезионного натяжения. После кавитации трахеида не может быть удалена эмболия и возвращена в рабочее состояние (за исключением нескольких продвинутых покрытосеменных, которые разработали такой механизм). Поэтому растениям стоит избегать кавитации. По этой причине ямки в стенках трахеиды имеют очень малый диаметр, чтобы предотвратить проникновение воздуха и образование пузырьков. Циклы замораживания-оттаивания являются основной причиной кавитации. Повреждение стенки трахеиды почти неизбежно приводит к просачиванию воздуха и кавитации, поэтому важно, чтобы многие трахеиды работали параллельно.
Кавитации трудно избежать, но как только она произошла, у растений есть ряд механизмов для сдерживания повреждение. Небольшие ямки соединяют соседние каналы, позволяя жидкости течь между ними, но не воздуху - хотя по иронии судьбы эти ямки, которые предотвращают распространение эмболий, также являются их основной причиной. Эти ямчатые поверхности дополнительно уменьшают поток воды через ксилему на 30%. Хвойные породы юрского периода разработали гениальное усовершенствование, используя клапанные структуры для изоляции кавитированных элементов. Эти структуры тор-марго имеют каплю, плавающую в середине пончика; когда одна сторона сбрасывает давление, капля засасывается в тор и блокирует дальнейший поток. Другие растения просто принимают кавитацию; например, в начале каждой весны у дубов вырастает кольцо из широких сосудов, ни одна из которых не выдерживает зимних заморозков. Каждую весну клены используют давление корней, чтобы выдавливать сок вверх от корней, выдавливая любые пузырьки воздуха.
Рост до высоты также использовал другую черту трахеид - поддержку, обеспечиваемую их одревесневшими стенками. Отмершие трахеиды были сохранены, чтобы сформировать прочный древесный стебель, в большинстве случаев производимый вторичной ксилемой. Однако у ранних растений трахеиды были слишком механически уязвимы и сохраняли центральное положение со слоем жесткой склеренхимы на внешнем крае стеблей. Даже когда трахеиды действительно играют структурную роль, они поддерживаются склеренхимной тканью.
Трахеиды заканчиваются стенками, которые оказывают большое сопротивление потоку; Элементы емкости имеют перфорированные торцевые стенки и расположены последовательно для работы, как если бы они были одним непрерывным резервуаром. Функция торцевых стенок, которые были стандартным состоянием в девоне, вероятно, заключалась в предотвращении эмболий. Эмболия - это образование воздушного пузыря в трахеиде. Это может произойти в результате замерзания или растворения газов из раствора. Как только образовалась эмболия, ее обычно нельзя удалить (но см. Ниже); пораженная клетка не может поднимать воду и становится бесполезной.
За исключением торцевых стенок, трахеиды предваскулярных растений могли работать при той же гидравлической проводимости, что и у первого сосудистого растения, Cooksonia.
Размер трахеид ограничен, поскольку они включают одиночная ячейка; это ограничивает их длину, что, в свою очередь, ограничивает их максимальный полезный диаметр до 80 мкм. Электропроводность растет в четвертой степени диаметра, поэтому увеличение диаметра имеет огромные преимущества; элементы сосуда, состоящие из ряда ячеек, соединенных на концах, преодолели этот предел и позволили сформироваться более крупным трубкам, достигающим диаметра до 500 мкм и длины до 10 м.
Сосуды впервые появились во время засушливых периодов с низким CO. 2 в поздней перми, в хвощах, папоротниках и Selaginellales независимо друг от друга, а позже появились в середине мелового периода у покрытосеменных растений и гнетофитов. Сосуды позволяют древесине той же площади поперечного сечения транспортировать примерно в сто раз больше воды, чем трахеиды! Это позволило растениям заполнить больше стеблей структурными волокнами, а также открыло новую нишу для виноградных лоз, которые могли транспортировать воду, не будучи такими же толстыми, как дерево, на котором они росли. Несмотря на эти преимущества, древесина на основе трахеид намного легче и, следовательно, дешевле в производстве, так как сосуды должны быть значительно усилены, чтобы избежать кавитации.
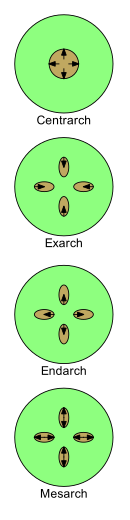 Модели развития ксилемы: ксилема в коричневом цвете; стрелки показывают направление развития от протоксилемы к метаксилеме.
Модели развития ксилемы: ксилема в коричневом цвете; стрелки показывают направление развития от протоксилемы к метаксилеме. Развитие ксилемы можно описать четырьмя терминами: центральный, экзарх, эндарх и мезарх. По мере развития у молодых растений его природа меняется от протоксилемы к метаксилеме (то есть от первой ксилемы к после ксилемы). Паттерны, в которых расположены протоксилема и метаксилема, важны при изучении морфологии растений.
По мере роста молодого сосудистого растения в его стеблях и корнях формируются одна или несколько нитей первичной ксилемы. Первая развивающаяся ксилема называется «протоксилема». По внешнему виду протоксилема обычно отличается более узкими сосудами, образованными более мелкими клетками. Некоторые из этих клеток имеют стенки, содержащие утолщения в виде колец или спиралей. Функционально протоксилема может расширяться: клетки могут увеличиваться в размерах и развиваться, в то время как стебель или корень удлиняются. Позже «метаксилема» развивается в тяжах ксилемы. Сосуды и клетки метаксилемы обычно больше; ячейки имеют утолщения, которые обычно имеют форму лестничных поперечных стержней (скалярная форма) или сплошных листов, за исключением отверстий или ямок (с ямками). Функционально метаксилема завершает свое развитие после того, как прекращается удлинение, когда клетки больше не нуждаются в увеличении размера.
Есть четыре основных образца расположения протоксилемы и метаксилемы в стебли и корни.
Остальные три термина используются там, где имеется более одной цепи первичной ксилемы.
В своей книге De plantis libri XVI (О растениях, в 16 книгах) (1583) итальянский врач и ботаник Андреа Чезальпино предположил, что растения вытягивают воду из почвы не за счет магнетизма (ut magnes ferrum trahit, поскольку магнитное железо притягивает) или за счет всасывания (вакуума), а путем поглощения, как это происходит в случае льняной ткани, губок., или порошки. Итальянский биолог Марчелло Мальпиги был первым человеком, который описал и проиллюстрировал сосуды ксилемы, что он и сделал в своей книге Anatome plantarum... (1675). Хотя Мальпиги считал, что ксилема содержит только воздух, британский врач и ботаник Нехемия Грю, который был современником Мальпиги, полагал, что сок поднимается как через кору, так и через ксилему. Однако, согласно Грю, капиллярное действие в ксилеме поднимет сок всего на несколько дюймов; Чтобы поднять сок на верхушку дерева, Грю предположил, что паренхимные клетки становятся набухшими и, таким образом, не только сжимают сок в трахеидах, но и выталкивают некоторое количество сока из паренхимы в трахеиды. В 1727 году английский священнослужитель и ботаник Стивен Хейлз показал, что транспирация листьями растения заставляет воду проходить через его ксилему. К 1891 году польско-немецкий ботаник Эдуард Страсбургер показал, что перенос воды в растениях не требует, чтобы клетки ксилемы были живыми.