
Войти
| Oogonium | |
|---|---|
| Идентификаторы | |
| MeSH | D009867 |
| FMA | 83673 |
| Анатомическая терминология [править в Викиданных ] | |
оогониум (множественное число оогония ) - это небольшая диплоидная клетка, которая при созревании образует приморд Иальный фолликул у женского плода или женского пола (гаплоидного или диплоидного) гаметангиума некоторых таллофитов.
оогонии образуются в большом количестве в результате митоза на ранних стадиях развития плода из первичные половые клетки. У людей они начинают развиваться между 4 и 8 неделями и присутствуют у плода между 5 и 30 неделями.
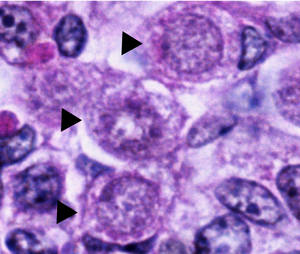 Окрашивание гематоксилином и эозином срезов гонад человека на E16.5. Спящие оогонии GO / G1 обозначены стрелками.
Окрашивание гематоксилином и эозином срезов гонад человека на E16.5. Спящие оогонии GO / G1 обозначены стрелками. Нормальные оогонии в человеческих яичниках имеют сферическую или яйцевидную форму и обнаруживаются среди соседних соматических клеток и ооцитов на разных фазах развития. Оогонии можно отличить от соседних соматических клеток под электронным микроскопом, наблюдая за их ядрами. Оогониальные ядра содержат беспорядочно рассредоточенный фибриллярный и зернистый материал, тогда как соматические клетки имеют более конденсированное ядро, которое создает более темный контур под микроскопом. Оогониальные ядра также содержат плотные выступающие ядрышки. хромосомный материал в ядре митотически делящихся оогоний выглядит как плотная масса, окруженная пузырьками или двойными мембранами.
цитоплазма оогониев похож на окружающие соматические клетки и аналогично содержит большие круглые митохондрии с боковыми кристами. Endoplasmic Reticulum (E.R.) оогоний, однако, очень недоразвит и состоит из нескольких небольших пузырьков. Некоторые из этих маленьких пузырьков содержат цистерны с рибосомами и находятся рядом с аппаратом Гольджи.
. Подвергающиеся дегенерации оогонии выглядят несколько иначе под электронным микроскопом. В этих оогониях хромосомы слипаются в неразличимую массу внутри ядра, а митохондрии и E.R., по-видимому, раздуты и разрушены. Дегенеративные оогонии обычно обнаруживаются частично или полностью поглощенными соседними соматическими клетками, что определяет фагоцитоз как способ устранения.
В бластоцисте эмбриона млекопитающего, примордиальные зародышевые клетки возникают из проксимальных эпибластов под влиянием внеэмбриональных сигналов. Эти половые клетки затем перемещаются посредством амебоидного движения к генитальному гребню и в конечном итоге в недифференцированные гонады плода. На 4-5 неделе развития гонады начинают дифференцироваться. В отсутствие Y-хромосомы гонады дифференцируются в яичники. По мере дифференциации яичников развиваются врастания, называемые корковыми канатиками. Здесь собираются первичные половые клетки.
В течение 6-8 недель эмбрионального развития самки (XX) первичные половые клетки растут и начинают дифференцироваться в оогонии. Оогонии пролиферируют посредством митоза в течение 9–22 недель эмбрионального развития. К 8-й неделе развития может быть до 600 000 оогоний и до 7 000 000 к 5-му месяцу.
В конце концов оогонии либо дегенерируют, либо далее дифференцируются в первичные ооциты посредством асимметричного деления. Асимметричное деление - это процесс митоза, в котором один оогоний делится неравномерно с образованием одной дочерней клетки, которая в конечном итоге станет ооцитом в процессе оогенеза, и одной дочерней клетки, которая является идентичным оогонию родительской клетке. Это происходит в период с 15-й по 7-й месяц эмбрионального развития. Большинство оогоний либо дегенерировали, либо дифференцировались в первичные ооциты при рождении.
Первичные ооциты подвергаются оогенезу, в котором они вступают в мейоз. Однако первичные ооциты арестовываются в профазе 1 первого мейоза и остаются в этой арестованной стадии до начала полового созревания у взрослых самок. Это контрастирует с первичными мужскими зародышевыми клетками, которые задерживаются на сперматогониальной стадии при рождении и не участвуют в сперматогенезе и мейозе с образованием первичных сперматоцитов до полового созревания у взрослого мужчины <. 32>
Регуляция и дифференцировка зародышевых клеток в первичные гаметоциты в конечном итоге зависит от пола эмбриона и дифференцировки гонад. У самок мышей белок RSPO1 отвечает за дифференциацию женских (XX) гонад в яичники. RSPO1 активирует сигнальный путь β-катенин путем активации Wnt4, что является важным этапом дифференцировки яичников. Исследования показали, что в яичниках, лишенных Rspo1 или Wnt4, будет наблюдаться изменение пола гонад, образование яйцеводов и дифференцировка соматических клеток Сертоли, которые способствуют развитию сперматозоидов.
после женского (XX) половые клетки собираются в недифференцированных половых железах, повышающая регуляция Stra8 необходима для дифференцировки зародышевых клеток в оогоний и, в конечном итоге, для вступления в мейоз. Одним из основных факторов, который способствует повышению регуляции Stra8, является инициация пути передачи сигналов β-катенина через RSPO1, который также отвечает за дифференцировку яичников. Поскольку RSPO1 продуцируется в соматических клетках, этот белок действует на половые клетки в паракринном режиме . Однако Rspo1 - не единственный фактор в регуляции Stra8. Многие другие факторы изучаются, и этот процесс все еще оценивается.
Теоретически оогонии либо дегенерируют, либо дифференцируются в первичные ооциты, которые попадают в оогенез и останавливаются в профазе I первого мейоза послеродового периода. Следовательно, считается, что у взрослых самок млекопитающих отсутствует популяция половых клеток, которые могут обновляться или регенерироваться, и вместо этого они имеют большую популяцию первичных ооцитов, которые задерживаются в первом мейозе до половой зрелости. В период полового созревания один первичный ооцит будет продолжать мейоз каждый менструальный цикл. Поскольку у человека отсутствуют регенерирующие половые клетки и оогонии, количество первичных ооцитов сокращается после каждого менструального цикла до менопаузы, когда у самки больше нет популяции первичных ооцитов.
Однако недавние исследования показали, что возобновляемые оогонии могут присутствовать в слизистой оболочке яичников самок людей, приматов и мышей. Считается, что эти половые клетки могут быть необходимы для поддержания репродуктивных фолликулов и развития ооцитов в зрелом возрасте. Также было обнаружено, что некоторые стволовые клетки могут мигрировать из костного мозга в яичники в качестве источника экстрагениальных половых клеток. Эти митотически активные половые клетки, обнаруженные у взрослых млекопитающих, были идентифицированы путем отслеживания нескольких маркеров, которые были обычными в ооцитах. Эти потенциально возобновляемые половые клетки были идентифицированы как положительные по этим важным маркерам ооцитов.
Обнаружение этих активных половых клеток и оогоний у взрослой женщины может быть очень полезным для продвижения исследований фертильности и лечения бесплодия. Зародышевые клетки были извлечены, изолированы и успешно выращены in vitro. Эти половые клетки использовались для восстановления фертильности у мышей, способствуя образованию и содержанию фолликулов у ранее бесплодных мышей. Также проводятся исследования возможной регенерации зародышевой линии у приматов. Митотически активные женские половые клетки человека могут оказаться очень полезными для нового метода развития эмбриональных стволовых клеток, который включает перенос ядра в зиготу. Использование этих функциональных оогоний может помочь в создании линий стволовых клеток, специфичных для пациента, с использованием этого метода.
Существуют значительные разногласия относительно существования оогониальных стволовых клеток млекопитающих. Разногласия заключаются в отрицательных данных, полученных во многих лабораториях США. Множественные подходы к проверке существования оогониальных стволовых клеток дали отрицательные результаты, и ни одна исследовательская группа в США не смогла воспроизвести первоначальные результаты.
 A Thalassiosira pseudonana diatom oogonium начало расширяться через клеточную стенку. Искусственная окраска обозначает хлорофилл (синий) и ДНК (красный).
A Thalassiosira pseudonana diatom oogonium начало расширяться через клеточную стенку. Искусственная окраска обозначает хлорофилл (синий) и ДНК (красный). В психологии и микологии, оогониум относится к женскому гаметангиуму если в этой структуре происходит объединение мужской (подвижной или неподвижной) и женской гаметы.
У Oomycota и некоторых других организмов женские оогонии и мужские эквиваленты антеридии являются результатом полового спороношения, то есть развития структур, внутри которых будет происходить мейоз. Гаплоидные ядра (гаметы) образуются в результате мейоза в антеридиях и оогониях, и когда происходит оплодотворение, образуется диплоидная ооспора, которая в конечном итоге прорастает на диплоидной соматической стадии жизненного цикла таллофита.
У многих водорослей (например, Chara ) основное растение гаплоидное; оогонии и антеридии образуют и продуцируют гаплоидные гаметы. Единственной диплоидной частью жизненного цикла является спора (оплодотворенная яйцеклетка), которая подвергается мейозу с образованием гаплоидных клеток, которые развиваются в новые растения. Это гаплонтический жизненный цикл (с зиготическим мейозом ).
Оогонии некоторых видов таллофитов обычно круглые или яйцевидные, с содержимым, разделенным на несколько одноядерных оосфер. Это отличается от мужских антеридий, которые имеют удлиненную форму и содержат несколько ядер.
У гетероталлических видов оогонии и антеридии расположены на гифальных ветвях различных таллофитов. колонии. Оогонии этого вида могут быть оплодотворены только антеридиями из другой колонии, поэтому самооплодотворение невозможно. Напротив, гомоталлические виды отображают оогонии и антеридии либо на одной и той же гифальной ветви, либо на отдельных гифальных ветвях, но в пределах одной и той же колонии.
В общем режиме При оплодотворении, обнаруженном у некоторых видов таллофитов, антеридии связываются с оогониями. Затем антеридии образуют трубки для оплодотворения, соединяющие цитоплазму антеридий с каждой оосферой внутри оогонии. Гаплоидное ядро (гамета) антеридия затем переносится через трубку для оплодотворения в оосферу и сливается с гаплоидным ядром оосферы, образуя диплоидную ооспору. Затем ооспора готова прорасти и превратиться во взрослую диплоидную соматическую стадию.
{kind=link}