
Войти
| Каталитический интрон группы I | |
|---|---|
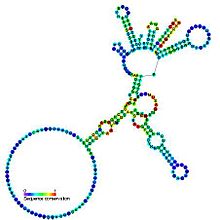 Предсказанная вторичная структура и сохранение последовательности каталитического интрона группы I Предсказанная вторичная структура и сохранение последовательности каталитического интрона группы I | |
| Идентификаторы | |
| Символ | Intron_gpI |
| Rfam | RF00028 |
| Другие данные | |
| РНК тип | Интрон |
| Домен ( s) | Eukaryota ; Бактерии ; Вирусы |
| GO | 0000372 |
| SO | 0000587 |
| PDB структуры | PDBe |
интроны группы I - это большие самосплайсирующие рибозимы. Они катализируют собственное вырезание из мРНК, тРНК и рРНК предшественников в широком диапазоне организмов. Ядро вторичной структуры состоит из девяти парных областей (P1-P9). Они сворачиваются по существу в два домена - домен P4-P6 (сформированный из стэкинга спиралей P5, P4, P6 и P6a) и домен P3-P9 (образованный из P8, P3, P7 и P9 спирали). Разметка вторичной структуры для этого семейства представляет только это консервативное ядро. Группа I интроны часто имеют длинные открытые рамки считывания, вставленные в области петли.
Сплайсинг интронов группы I обрабатывается двумя последовательные реакции переэтерификации. экзогенный гуанозин или гуанозин нуклеотид (exoG) сначала стыкуется с активным G-связывающим сайтом, расположенным в P7, и его 3'-OH выравнивается для атаки фосфодиэфирная связь в 5'-сайте сплайсинга, расположенном в P1, в результате чего образуется свободная 3'-OH группа в верхнем экзоне и exoG прикрепляется к 5'-концу интрон. Затем конец G (омега G) интрона меняет местами exoG и занимает сайт связывания G, чтобы организовать вторую реакцию переноса сложного эфира: 3'-OH группа вышестоящего экзона в P1 выравнивается для атаки 3'-сплайсинга. сайт в P10, приводящий к лигированию соседних вышестоящих и нижележащих экзонов и высвобождению каталитического интрона.

Механизм двух ионов металлов, наблюдаемый в протеин полимеразах и фосфатазах, был предложен для использования интронами группы I и группы II для процесса переноса фосфорила реакции, что было однозначно доказано структурой высокого разрешения интрона Azoarcus группы I в 2006 году.
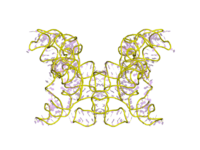 Трехмерное изображение каталитического интрона группы I. На этом изображении показан активный центр в кристаллической структуре рибозима Tetrahymena.
Трехмерное изображение каталитического интрона группы I. На этом изображении показан активный центр в кристаллической структуре рибозима Tetrahymena.  Трехмерное изображение каталитического интрона группы I. Это кристаллическая структура комплекса рибозим-продукт фага Twort группы I.
Трехмерное изображение каталитического интрона группы I. Это кристаллическая структура комплекса рибозим-продукт фага Twort группы I.  Трехмерное изображение каталитического интрона группы I. Это структура рибозима Tetrahymena с базовым тройным сэндвичем и ионом металла в активном центре.
Трехмерное изображение каталитического интрона группы I. Это структура рибозима Tetrahymena с базовым тройным сэндвичем и ионом металла в активном центре. С начала 1990-х годов ученые начали изучать, как интрон группы I достигает своей естественной структуры. in vitro и некоторые механизмы сворачивания РНК к настоящему времени оценены. Принято считать, что третичная структура складывается после образования вторичной структуры. Во время сворачивания молекулы РНК быстро заселяются в различные промежуточные соединения сворачивания, промежуточные соединения, содержащие нативные взаимодействия, далее сворачиваются в нативную структуру посредством быстрого пути сворачивания, в то время как молекулы, содержащие ненативные взаимодействия, захватываются метастабильными или стабильными неродные конформации, и процесс преобразования в нативную структуру происходит очень медленно. Очевидно, что интроны группы I, различающиеся набором периферических элементов, проявляют разные потенциалы вхождения в путь быстрого сворачивания. Между тем, совместная сборка третичной структуры важна для сворачивания собственной структуры. Тем не менее, сворачивание интронов группы I in vitro сталкивается как с термодинамическими, так и с кинетическими проблемами. Было показано, что некоторые связывающие РНК белки и шапероны способствуют сворачиванию интронов группы I in vitro и в бактериях за счет стабилизации нативных промежуточных продуктов и дестабилизации ненативных структур, соответственно.
Интроны группы I распределены у бактерий, низших эукариот и высших растений. Однако их появление у бактерий кажется более спорадическим, чем у низших эукариот, и они стали преобладать у высших растений. Гены , которые прерывают интроны группы I, существенно различаются: они прерывают гены рРНК, мРНК и тРНК в бактериальных геномах, а также в геномах митохондриальный и хлоропластный геномы низших эукариот, но вторгаются только в гены рРНК в ядерном геноме низших эукариот. У высших растений эти интроны, по-видимому, ограничены несколькими генами тРНК и мРНК хлоропластов и митохондрий.
Интроны группы I также обнаруживаются встроенными в гены большого разнообразия бактериофагов грамположительных бактерий. Однако их распространение в фаге грамотрицательных бактерий в основном ограничивается бактериофагами Т4, Т-четный и Т7-подобный.
Обе теории интрона-раннего и интрон-позднего нашли доказательства в объяснении происхождения интронов группы I. Некоторые интроны группы I кодируют самонаводящуюся эндонуклеазу (HEG), которая катализирует подвижность интронов. Предполагается, что HEG перемещают интрон из одного места в другое, из одного организма в другой и, таким образом, объясняют широкое распространение эгоистичных интронов группы I. Никакой биологической роли интронов группы I до сих пор не выявлено, за исключением сплайсинга самих себя из предшественника, чтобы предотвратить смерть хозяина, которым они живут. Также обнаружено, что небольшое количество интронов группы I кодирует класс белков, называемых матуразами, которые облегчают сплайсинг интронов.