
Войти
| ДНК-полимераза II | |||||||
|---|---|---|---|---|---|---|---|
 Кристаллическая структура ДНК pol II (на основе записи PDB 3K5M) Кристаллическая структура ДНК pol II (на основе записи PDB 3K5M) | |||||||
| Идентификаторы | |||||||
| Организм | Escherichia coli. (ул. К-12 подл. MG1655) | ||||||
| Symbol | polB | ||||||
| Entrez | 944779 | ||||||
| PDB | 3K5M | ||||||
| RefSeq (Prot) | NP_414602.1 | ||||||
| UniProt | P21189 | ||||||
| Другие данные | |||||||
| Номер EC | 2.7.7.7 | ||||||
| Хромосома | геном: 0,06 - 0,07 Мб | ||||||
| |||||||
ДНК-полимераза II (также известная как ДНК Pol II или Pol II ) представляет собой прокариотическую ДНК-зависимую ДНК полимераза, кодируемая геном PolB.
ДНК-полимераза II представляет собой белок массой 89,9 кДа и является членом семейства ДНК-полимераз B. Первоначально он был изолирован Томасом Корнбергом в 1970 году и охарактеризован в течение следующих нескольких лет. Функциональность Pol II in vivo является предметом дискуссий, однако общее мнение показывает, что Pol II в первую очередь участвует в качестве резервного фермента в репликации прокариотической ДНК. Фермент обладает способностью к синтезу ДНК 5 '→ 3' , а также активностью проверки 3 '→ 5' экзонуклеазы. ДНК Pol II взаимодействует с множеством партнеров по связыванию, общих с ДНК Pol III, чтобы повысить ее точность и процессивность.
ДНК-полимераза I была первой ДНК-направленной ДНК-полимеразой, выделенной из E. coli. Несколько исследований с участием этого изолированного фермента показали, что ДНК pol I, скорее всего, участвует в репарационной репликации и не является основной репликативной полимеразой. Чтобы лучше понять in vivo роль ДНК pol I, в 1969 году Де Люсия и Кэрнс создали мутанты E. coli , дефицитные по этому ферменту (названные Pol A1). Как было охарактеризовано, этот новый мутантный штамм был более чувствителен к ультрафиолетовому свету, что подтверждает гипотезу о том, что ДНК pol I участвует в репарационной репликации. Мутант рос с той же скоростью, что и дикого типа, что указывает на присутствие другого фермента, ответственного за репликацию ДНК. Затем последовали выделение и характеристика этой новой полимеразы, участвующей в полуконсервативной репликации ДНК, в параллельных исследованиях, проведенных в нескольких лабораториях. Новая полимераза была названа ДНК-полимеразой II и какое-то время считалась основным репликативным ферментом E. coli. ДНК pol II была впервые кристаллизована Anderson et al. в 1994.
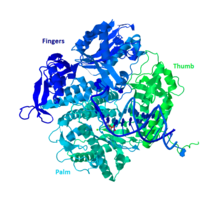
ДНК Pol II представляет собой белок 89,9 кДа, состоящий из 783 аминокислот, который кодируется геном polB (dinA). Глобулярный белок, ДНК Pol II функционирует как мономер, тогда как многие другие полимеразы образуют комплексы. Есть три основных части этого мономера, которые в просторечии называются ладонью, пальцами и большим пальцем. Эта «рука» смыкается вокруг нити ДНК. На ладони комплекса три каталитических остатка, которые будут координироваться с двумя ионами двухвалентных металлов, чтобы функционировать. ДНК Pol II имеет большое количество копий в клетке, около 30-50, тогда как уровень ДНК Pol III в клетке в пять раз меньше.
 Общая «ручная» структура полимеразы группы B (на основе записи PDB 3NCI)
Общая «ручная» структура полимеразы группы B (на основе записи PDB 3NCI) Большинство полимераз сгруппированы в семейства на основе аналогичной структуры и функции. ДНК Pol II попадает в группу B вместе с человеческой ДНК Pol α, δ, ϵ и ζ. Все это гомологи RB69, 9 ° N-7 и Tgo. Другие члены группы B имеют по крайней мере одну другую субъединицу, которая делает ДНК Pol II уникальной.
Полимеразы все участвуют в ДНК репликация в некотором качестве, синтезирующая цепочки нуклеиновых кислот. Репликация ДНК - жизненно важный аспект размножения клетки. Без репликации своей ДНК клетка не может делиться и делиться своей генетической информацией с потомством. У прокариот, таких как E. coli, ДНК Pol III является основной полимеразой, участвующей в репликации ДНК. Хотя ДНК Pol II не является основным фактором репликации хромосом, у нее есть и другие роли.
ДНК Pol II действительно участвует в репликации ДНК. Хотя он может быть не таким быстрым, как ДНК Pol III, у него есть некоторые способности, которые делают его эффективным ферментом. Этот фермент обладает ассоциированной активностью 3 '→ 5' экзонуклеазы наряду с активностью примазы. ДНК Pol II представляет собой фермент высокой точности с частотой ошибок замены ≤ 2 × 10 и частотой ошибок сдвига рамки -1 ≤ 1 × 10. DNA Pol II может корректировать и обрабатывать несоответствия, вызванные Pol III. Banach-Orlowska et al. показали, что ДНК Pol II участвует в репликации, но она зависит от цепи и предпочтительно реплицирует отстающую цепь. Предлагаемый механизм предполагает, что когда ДНК Pol III останавливается или становится нефункциональной, тогда ДНК Pol II может быть специфически задействована в точке репликации и продолжить репликацию.
Существует много разных способов повреждения ДНК, от УФ-повреждений до окисления, поэтому логично, что существуют разные типы полимераз для устранения этих повреждений. Одна из важных ролей ДНК Pol II - это основная полимераза для восстановления межцепочечных поперечных связей. Межцепочечные сшивки вызываются такими химическими веществами, как азотистый иприт и псорален, которые вызывают цитотоксические поражения. Восстановить эти повреждения сложно, потому что обе цепи ДНК были повреждены химическим агентом, и, следовательно, генетическая информация на обеих цепях неверна. Точный механизм фиксации этих межнитевых поперечных связей все еще исследуется, но известно, что Pol II принимает активное участие.
ДНК Pol II не является наиболее изученной полимеразой поэтому есть много предполагаемых функций этого фермента, которые все являются вероятными функциями, но в конечном итоге не подтверждены:
 Ремонтный активный сайт ДНК-полимеразы II (PDB ID: 3K5M)
Ремонтный активный сайт ДНК-полимеразы II (PDB ID: 3K5M) Во время репликации ДНК пары оснований в последовательности подвержены повреждению. Поврежденная последовательность ДНК может вызвать остановку репликации. Чтобы исправить ошибку в последовательности, ДНК Pol II катализирует восстановление пар нуклеотидных оснований. Исследования in vitro показали, что Pol II иногда взаимодействует с дополнительными белками Pol III (β-Clamp и комплекс загрузки Clamp), обеспечивая доступ Pol II к растущей зарождающейся цепи. Что касается функции ДНК Pol II во время репликации ДНК, это имеет смысл, поскольку любые ошибки, которые производит Pol III, будут происходить в растущей цепи, а не в консервативной цепи. N-концевой домен ДНК Pol II отвечает за ассоциацию и диссоциацию цепи ДНК с каталитической субъединицей. Скорее всего, есть два сайта в N-концевом домене ДНК Pol II, которые распознают одноцепочечную ДНК. Один (е) сайт (ы) отвечает за привлечение одноцепочечной ДНК к ДНК Pol II, а другой сайт (ы) отвечает за диссоциацию одноцепочечной ДНК от ДНК Pol II.
 Активный сайт ДНК Pol II (PDB ID 3K5M))
Активный сайт ДНК Pol II (PDB ID 3K5M)) При связывании субстрата ДНК Pol II связывает нуклеозидтрифосфаты для поддержания структуры ДНК с водородными связями. Затем связывается правильный dNTP, и ферментный комплекс претерпевает конформационные изменения субдоменов и аминокислотных остатков. Эти конформационные изменения позволяют ускорить синтез репарации. Активный центр содержит два иона Mg, которые стабилизируются каталитическими аспарагиновыми кислотами D419 и D547. Ионы магния связываются с ДНК вместе с дНТФ в открытом состоянии и координируют конформационные изменения аминокислотных остатков активного центра для проведения катализа (закрытое состояние). После высвобождения ионов магния фермент возвращается в открытое состояние.
ДНК-полимераза II является членом семейства полимераз B и поддерживает полимеразу III при репликации ДНК перемещается от 3'-конца к 5'-концу. В случае, когда Полимераза III останавливается во время ошибки репликации, Полимераза II может прервать и вырезать несовпадающие основания. Полимераза II имеет гораздо более высокий коэффициент точности, чем Полимераза III, а это означает, что вероятность возникновения неправильного спаривания гораздо меньше. Без этапа корректирующего считывания Полимеразы II Полимераза III расширит ошибочные спаривания и, таким образом, создаст мутацию.
Помимо защиты от мутаций, которые могут быть вызваны Полимеразой III, Полимераза II действует для защиты от мутаций, вызванных Полимеразой IV. Полимераза IV гораздо более подвержена ошибкам, чем Полимераза II, но также функционирует для восстановления несовпадающих пар оснований, начиная с 3'-конца. Полимераза II защищает 3'-конец от полимеразы IV и блокирует ее действие. Эта защита предотвратит образование мутаций, пока Полимераза II функционирует нормально. Если Полимераза II будет выведена из строя в результате мутации или отключена другими факторами, Полимераза IV займет ее место, чтобы исправить ошибочно спаренные основания.
В то время как Полимераза II не будет нормально функционировать вместе с эукариотическими членами семейства B, у него действительно сходные структурные и функциональные мотивы. Члены семейства B включают полимеразу α, ε, ζ и δ. Все эти полимеразы предназначены для коррекции вновь синтезированной ДНК в направлении 3 '→ 5'. Эти полимеразы способны синтезировать ДНК как на ведущих, так и на отстающих цепях. Этот класс полимеразы имеет тенденцию быть очень точным, что позволяет им исправлять любые нарушения спаривания, возникающие во время синтеза ДНК.
ДНК-полимераза II естественным образом присутствует в клетке, что обычно составляет пять раз больше, чем количество Полимеразы III. Это большее количество позволяет Полимеразе II превосходить Полимеразу III в случае неправильного спаривания. Это количество может быть увеличено после индукции SOS-ответа, который активирует ген polB, поэтому количество полимеразы II увеличивается примерно в семь раз. Хотя Полимераза II может работать с обеими цепями, было показано, что она предпочитает отстающую цепь, а не ведущую.