Войти
Филогенетические сравнительные методы (PCM ) используют информацию об исторических взаимосвязях родословных (филогении ) для проверки эволюционных гипотез. Сравнительный метод имеет долгую историю в эволюционной биологии; действительно, Чарльз Дарвин использовал различия и сходства между видами в качестве основного источника доказательств в Происхождении видов. Однако тот факт, что близкородственные линии имеют много общих черт и комбинаций черт в результате процесса происхождения с модификацией, означает, что линии не являются независимыми. Это осознание вдохновило на разработку явно филогенетических сравнительных методов. Первоначально эти методы были разработаны в основном для контроля филогенетического анамнеза при тестировании на адаптацию ; однако в последние годы использование этого термина расширилось и теперь включает любое использование филогении в статистических тестах. Хотя большинство исследований, в которых используются ПКМ, сосредоточены на современных организмах, многие методы также могут применяться к вымершим таксонам и могут включать информацию из летописи окаменелостей.
ПКМ, как правило, можно разделить на два типа подходов: те, которые предполагают эволюционный история какого-либо признака (фенотип или генетический ) на протяжении филогении и тех, которые определяют сам процесс эволюционного ветвления (коэффициенты диверсификации ), хотя есть некоторые подходы которые делают и то, и другое одновременно. Обычно дерево, которое используется вместе с PCM, оценивается независимо (см. вычислительная филогенетика ), так что предполагается, что известны как отношения между линиями, так и длина разделяющих их ветвей.
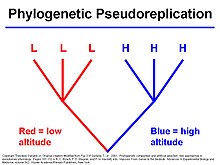
Филогенетические сравнительные подходы могут дополнять другие способы изучения адаптации, например, изучение природных популяций, экспериментальные исследования и математические модели. Межвидовые сравнения позволяют исследователям оценить общность эволюционных явлений, рассматривая независимые эволюционные события. Такой подход особенно полезен, когда внутри видов мало или совсем нет изменений. И поскольку их можно использовать для явного моделирования эволюционных процессов, происходящих в течение очень долгих периодов времени, они могут дать представление о макроэволюционных вопросах, которые когда-то были исключительной областью палеонтологии.
 ареала обитания 49 видов млекопитающие по отношению к своим. Виды с более крупным телом, как правило, имеют больший ареал обитания, но при любом заданном размере тела представители отряда Carnivora (плотоядные и всеядные ), как правило, имеют более крупный дом. диапазоны, чем копытные (все из которых травоядные ). Считается ли это различие статистически значимым, зависит от того, какой тип анализа применяется
ареала обитания 49 видов млекопитающие по отношению к своим. Виды с более крупным телом, как правило, имеют больший ареал обитания, но при любом заданном размере тела представители отряда Carnivora (плотоядные и всеядные ), как правило, имеют более крупный дом. диапазоны, чем копытные (все из которых травоядные ). Считается ли это различие статистически значимым, зависит от того, какой тип анализа применяется  семенников масса различных видов приматов по отношению к их системе и системе спаривания. Виды с более крупным телом, как правило, имеют более крупные семенники, но у видов с любым заданным размером тела, у которых самки, как правило, спариваются с несколькими самцами, имеют самцов с более крупными семенниками.
семенников масса различных видов приматов по отношению к их системе и системе спаривания. Виды с более крупным телом, как правило, имеют более крупные семенники, но у видов с любым заданным размером тела, у которых самки, как правило, спариваются с несколькими самцами, имеют самцов с более крупными семенниками. Филогенетические сравнительные методы обычно применяются к таким вопросам, как:
→ Пример: как масса мозга изменяется по отношению к массе тела ?
→ Пример: у псовых сердце больше, чем у кошачьих ?
→ Пример: имеют ли плотоядные животные более крупные зоны обитания чем травоядные?
→ Пример: где эндотермия возникла в линии, которая привела к млекопитающим?
→ Пример: где, когда и почему развились плаценты и живорождения ?
→ Пример: являются ли поведенческие черты более лабильными в процессе эволюции?
→ Пример: почему у мелких видов более короткая продолжительность жизни чем их более крупные родственники?
 Стандартизированные контрасты используются в обычных статистических процедурах с ограничением, что все регрессии, корреляции, анализ ковариации и т. Д., Должны проходить через источник.
Стандартизированные контрасты используются в обычных статистических процедурах с ограничением, что все регрессии, корреляции, анализ ковариации и т. Д., Должны проходить через источник. Фельзенштейн в 1985 году предложил первый общий статистический метод для включения филогенетической информации, т. Е. Первый, который мог использовать любую произвольную топологию (ветвление порядок) и заданный набор длин ветвей. Этот метод теперь распознается как алгоритм, который реализует частный случай так называемых филогенетических обобщенных моделей наименьших квадратов. логика метода заключается в использовании филогенетической информации (и предполагаемой модели эволюции признака, подобной броуновскому движению ) для преобразования исходных данных кончика (средние значения для набора видов) в значения, которые статистически независимы и одинаково распределены.
Алгоритм включает в себя вычисление значений на внутренних узлах в качестве промежуточного шага, но они обычно не используются для выводов сами по себе. Исключение составляет базальный (корневой) узел, который можно интерпретировать как оценку наследственного значения для всего дерева (при условии, что никакого направленного [например, правило Копа ] не произошло) или как филогенетически взвешенная оценка среднего для всего набора верхушечных видов (терминальных таксонов). Значение в корне эквивалентно значению, полученному с помощью алгоритма «экономия квадратов с изменением квадрата», а также является оценкой максимального правдоподобия при броуновском движении. Алгебра независимых контрастов также может использоваться для вычисления стандартной ошибки или доверительного интервала.
Вероятно, наиболее часто используемым PCM является филогенетический обобщенный наименьших квадратов (PGLS). Этот подход используется для проверки наличия связи между двумя (или более) переменными с учетом того факта, что происхождение не является независимым. Этот метод является частным случаем обобщенных наименьших квадратов (GLS), и поэтому оценка PGLS также несмещенная, согласованная, эффективная, и асимптотически нормальный. Во многих статистических ситуациях, когда используется GLS (или обычный метод наименьших квадратов [OLS]), предполагается, что остаточные ошибки ε являются независимыми и одинаково распределенными случайными величинами, которые считаются нормальными

, тогда как в PGLS предполагается, что ошибки распределяются как

где V - матрица ожидаемой дисперсии и ковариации остатков с учетом эволюционной модели и филогенетическое дерево. Таким образом, отображается структура остатков, а не сами переменные. Это уже давно вызывает путаницу в научной литературе. Для структуры V был предложен ряд моделей, таких как броуновское движение Орнштейн-Уленбек и λ-модель Пагеля. (Когда используется модель броуновского движения, PGLS идентичен независимой оценке контрастов.) В PGLS параметры эволюционной модели обычно оцениваются совместно с параметрами регрессии.
PGLS может применяться только к вопросам, где зависимая переменная непрерывно распределена; однако филогенетическое дерево также может быть включено в остаточное распределение обобщенных линейных моделей, что позволяет обобщить подход к более широкому набору распределений для ответа.
 Данные для непрерывно оцениваемого признака могут быть смоделированы таким образом, что таксоны на концах гипотетического филогенетического дерева будут демонстрировать филогенетический сигнал, т.е. близкородственные виды будут иметь тенденцию походить друг на друга.
Данные для непрерывно оцениваемого признака могут быть смоделированы таким образом, что таксоны на концах гипотетического филогенетического дерева будут демонстрировать филогенетический сигнал, т.е. близкородственные виды будут иметь тенденцию походить друг на друга. Мартинс и Гарланд предложили в 1991 году, что одним из способов учета филогенетических отношений при проведении статистического анализа было использование компьютерного моделирования для создания множества наборов данных, согласующихся с проверяемой нулевой гипотезой (например, отсутствие корреляции между двумя признаками, отсутствие разницы между двумя экологически определенные группы видов), но это имитирует эволюцию по соответствующему филогенетическому дереву. Если такие наборы данных (обычно 1000 или более) анализируются с помощью той же статистической процедуры, которая используется для анализа реального набора данных, то результаты для смоделированных наборов данных можно использовать для создания филогенетически правильных (или «ПК») нулевых распределений статистика теста (например, коэффициент корреляции, t, F). Такие подходы к моделированию также можно комбинировать с такими методами, как филогенетически независимые контрасты или PGLS (см. Выше).