
Войти
| Эпотилоны | |
|---|---|
 Эпотилоны A (R = H) и B (R = CH 3) | |
| Химические формулы | A:C26H39NO6S. B:C27H41NO6S |
| Молекулярные массы | A:493,66 г / моль. B:507,68 г / моль |
| Номера CAS | A:152044-53-6. B:152044-54-7 |
| PubChem | A:448799. B:448013 |
 Эпотилоны C (R = H) и D (R = CH 3) | |
| Химические формулы | C:C26H39NO5S. D:C27H41NO5S |
| Молекулярные массы | C:477,66 г / моль. D:491,68 г / моль |
| Номера CAS | D:189453-10-9 |
| PubChem | C:9891226. D:447865 |
 Эпотилоны E (R = H) и d F (R = CH 3) | |
| Химические формулы | E:C26H39NO7S. F:C27H41NO7S |
| Молекулярные массы | E:509,66 г / моль. F:523,68 г / моль |
| Заявление об ограничении ответственности и ссылки | |
эпотилоны представляют собой класс потенциальных лекарств от рака. Как и таксаны, они предотвращают деление раковых клеток, вмешиваясь в тубулин, но в ранних исследованиях эпотилоны обладают большей эффективностью и более мягкими побочными эффектами, чем таксаны.
По состоянию на сентябрь. 2008 г. были идентифицированы и охарактеризованы эпотилоны A - F . Ранние исследования линий раковых клеток и пациентов с раком человека показали более высокую эффективность, чем таксаны. Их механизм действия похож, но их химическая структура проще. Из-за их лучшей растворимости в воде кремофоры (солюбилизирующие агенты, используемые для паклитаксела, которые могут влиять на сердечную функцию и вызывать тяжелую гиперчувствительность) не нужны. Эндотоксиноподобные свойства, известные у паклитаксела, такие как активация макрофагов, синтезирующих воспалительные цитокины и оксид азота, не наблюдаются для эпотилона B.
Эпотилоны первоначально были идентифицированы как метаболиты, продуцируемые в почве. myxobacterium Sorangium cellulosum.
Структура эпотилона А была определена в 1996 году с помощью рентгеновской кристаллографии.
Основным механизмом класса эпотилонов является ингибирование функции микротрубочек. Микротрубочки необходимы для деления клеток, поэтому эпотилоны не позволяют клеткам делиться должным образом. Эпотилон B обладает теми же биологическими эффектами, что и паклитаксел, как in vitro, так и в культивируемых клетках. Это потому, что они имеют один и тот же сайт связывания, а также сродство связывания с микротрубочкой. Подобно паклитакселу, эпотилон B связывается с субъединицей гетеродимера αβ-тубулина. После связывания скорость диссоциации αβ-тубулина снижается, что стабилизирует микротрубочки. Кроме того, было показано, что эпотилон B вызывает полимеризацию тубулина в микротрубочки без присутствия GTP. Это вызвано образованием пучков микротрубочек по всей цитоплазме. Наконец, эпотилон B также вызывает остановку клеточного цикла в фазе перехода G2-M, что приводит к цитотоксичности и, в конечном итоге, к апоптозу клеток. Способность эпотилона подавлять функцию веретена обычно связывают с его подавлением динамики микротрубочек; но недавние исследования показали, что подавление динамики происходит при более низких концентрациях, чем те, которые необходимы для блокирования митоза. При более высоких концентрациях антимитотика паклитаксел, по-видимому, действует, подавляя отслоение микротрубочек от центросом, процесс, который обычно активируется во время митоза. Вполне возможно, что эпотилон также может действовать через аналогичный механизм.
Один аналог, иксабепилон, был одобрен в октябре 2007 года в США. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов для лечения агрессивного метастатического или местнораспространенного рака груди, больше не поддающегося лечению доступными в настоящее время химиотерапевтическими препаратами. В ноябре 2008 г. EMEA отказало в разрешении на продажу иксабепилона.
Несколько синтетических аналогов эпотилона в настоящее время проходят клиническую разработку для лечения различных раковых заболеваний.
Эпотилон B, как было доказано, обладает сильной противораковой активностью in vivo при допустимых уровнях доз на нескольких моделях ксенотрансплантатов человека. В результате, эпотилон B (патупилон) и его различные аналоги по состоянию на 2001 год проходят различные клинические фазы: патупилон и полностью синтетический сагопилон [SH-Y03757A, ZK-EPO, химическая структура ] находятся в фазе II испытаний. ; и BMS-247550 в фазе I испытаний).
Объявлены результаты исследования фазы III с иксабепилоном (BMS-247550) в сочетании с капецитабином при метастатическом раке молочной железы (2007 г. - привело к одобрению FDA).
Патупилон потерпел неудачу в испытании фазы III по раку яичников в 2010 году.
Утиделон - это генно-инженерный аналог эпотилона, который показал преимущества в исследовании III фазы рака груди, когда добавлен к капецитабину.
В связи с высокой эффективностью и клинической потребностью в лечении рака эпотилоны были мишенью для многих общих синтезов. Первой группой, опубликовавшей полный синтез эпотилонов, была S. J. Danishefsky et al. в 1996 г. Этот полный синтез эпотилона А был достигнут посредством внутримолекулярной конденсации енолят-альдегида сложного эфира. Другие синтезы эпотилонов были опубликованы Николау, Шинцером, Мульцером и Каррейрой. В этом подходе были сконструированы ключевые строительные блоки альдегид, глицидолы и кетокислота, которые были присоединены к предшественнику метатезиса олефинов посредством альдольной реакции и затем соединение этерификации. Катализатор Граббса использовали для закрытия бис-концевого олефина соединения-предшественника. Полученные соединения представляли собой цис- и тран-макроциклические изомеры с отдельными стереоцентрами. Эпоксидирование цис- и транс-олефинов дает эпотилон А и его аналоги.
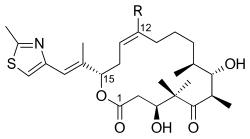
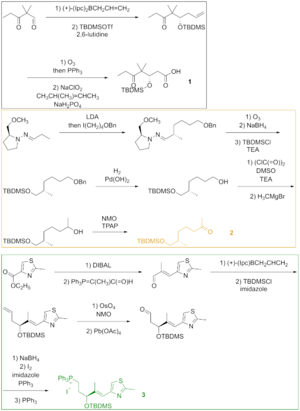
Один из полных синтезов эпотилона B описан ниже и был описан лабораторией K. К. Николау. Ретросинтетический анализ выявил 1, 2и 3 в качестве строительных блоков (рисунок 1).
 Фигура 1
Фигура 1 Как видно на Рисунке 2, кетокислота 1 была образована из кетоальдегида, который был преобразован в силиловый эфир посредством асимметричного аллилбората и силилирования. полученного спирта. Озонолиз простого силилового эфира и окисление альдегида по Линдгрену - Пинником с образованием кетокислоты. Кетон 2 был сконструирован посредством эндерс-алкилирования, исходя из гидразона. Озонолиз, последняя стадия алкилирования Эндера, сопровождалось восстановлением альдегида и силилированием образующегося спирта. Гидрогенолиз бензилового эфира давал спирт, который окисляли в условиях Сверна и алкилировали реагентом Гриньяра с получением вторичного спирта. Окисление этого спирта реагентом Лея-Гриффита давало желаемый кетон. Тиазол 3 синтезировали из сложного эфира, который восстанавливали с помощью диизобутилалюминийгидрида, и альдегид подвергали взаимодействию со стабилизированным илидом в реакции Виттига. Асимметричное аллилборирование α, β-ненасыщенного альдегида и защита гидроксигруппы давали силиловый эфир, концевой олефин которого реагировал с тетроксидом осмия до диола, который расщеплялся тетраацетатом свинца для получения альдегида. Восстановление, йодирование и обработка трифенилфосфином привели к соли фосфония.
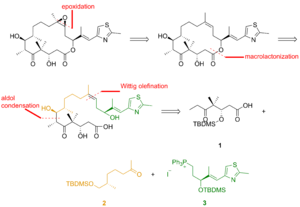
 Фиг. 2
Фиг. 2 Фрагменты 1, 2и 3 реагировали друг с другом для доставки эпотилона B в подходе, включающем реакцию Виттига, альдольную реакцию и этерификация Ямагути (Фиг.3). Для разделения диастереомеров использовали препаративную тонкослойную хроматографию.
 Рисунок 3
Рисунок 3 Эпотилон B представляет собой 16-членный поликетид макролактон с метиловой тиазольной группой, связанной с макроцикл олефиновой связью. Поликетидный скелет был синтезирован с помощью поликетидсинтазы (PKS) типа I, а тиазольное кольцо было получено из цистеина, включенного нерибосомальной пептидной синтетазой (NRPS). В этом биосинтезе и PKS, и NRPS используют белки-носители, которые посттрансляционно модифицированы группами фосфопантетеина, для присоединения к растущей цепи. PKS использует тиоэфир кофермента-A, чтобы катализировать реакцию и модифицировать субстраты путем селективного восстановления β-карбонила до гидроксила (кеторедуктаза, KR), алкена ( Дегидратаза, DH) и алкан (Эноилредуктаза, ER). PKS-I может также метилировать α-углерод субстрата. NRPS, с другой стороны, использует аминокислоты, активированные на ферменте в качестве аминоациладенилатов. В отличие от PKS, эпимеризация, N-метилирование и образование гетероцикла происходят в ферменте NRPS.
 Рисунок 4
Рисунок 4  Рисунок 5
Рисунок 5 Эпотилон B начинается с 2-метил-4-карбокситиазола., который был сформирован за счет трансляционной связи между модулем PKS, EPOS A (epoA) и NRPS, модулем EPOS P (epoP). EPOS A содержит модифицированную β-кетоацилсинтазу (малонил-ACP декарбоксилазу, KSQ), ацилтрансферазу (AT), еноилредуктазу (ER) и домен белка-носителя ацила (ACP). Однако EPOS P содержит домен гетероцилирования, аденилирования, оксидазы и тиолированный домен. Эти домены важны, потому что они участвуют в образовании пятичленного гетероциклического кольца тиазола. Как видно на фиг. 4, EPOS P активирует цистеин и связывает активированный цистеин как аминоацил-S-PCP. После связывания цистеина EPOS A загружает ацетатное звено в комплекс EPOS P, инициируя таким образом образование тиазолинового кольца за счет внутримолекулярной циклодегидратации.
Как только 2-метилтиазольное кольцо образовалось. был получен, он затем переносится на PKS EPOS B (epoB), EPOS C (epoC), EPOS D (epoD), EPOS E (epoE) и EPOS F (epoF) для последующего удлинения и модификации с образованием олефиновой связи., 16-членное кольцо и эпоксид, как показано на Фиг.5 . Следует отметить один важный момент - синтез гем-диметильного звена в модуле 7. Эти два диметила не были синтезированы двумя последовательными С-метилированием. Вместо этого одна из метильной группы была получена из пропионатного удлинителя, тогда как вторая метильная группа была интегрирована с помощью домена C-метилтрансферазы.
{kind=link}