
Войти
В эволюционной биологии предвзятость развития относится к продукции, направленной против или в сторону определенного онтогенетические траектории, которые в конечном итоге влияют на направление и исход эволюционных изменений, влияя на скорости, величины, направления и пределы признака эволюции. Исторически этот термин был синонимом ограничения развития, однако последнее время интерпретировалось как относящееся исключительно к отрицательной роли развития в эволюции.
 Эволюция происходит через изменения в развитии
Эволюция происходит через изменения в развитии В современной эволюционной биологии идея смещения развития сводится к встроен в поток мысли ок. lled Структурализм (также «интернализм» или «вариационный структурализм») сосредоточился на роли организма как причинной силы эволюционных изменений. С точки зрения структуралистов, фенотипическая эволюция является результатом действия естественного отбора на ранее «отфильтрованные» вариации в течение онтогенеза. Это контрастирует с взглядами функционалистов (также «адаптационистов», «пан-селекционистов» или «экстерналистов»), согласно которым фенотипическая эволюция является результатом только взаимодействия между детерминированным действием естественного отбора и изменчивостью, вызванной мутацией.
Обоснование роли организма или, в частности, эмбриона как причинной силы в эволюции и существования предвзятости, заключается в следующем: традиционный неодарвинистский подход к объяснению процесса, лежащего в основе эволюционное изменение - это естественный отбор, воздействующий на наследственные вариации, вызванные генетическими мутациями. Однако естественный отбор действует на фенотипы, и мутация сама по себе не вызывает фенотипических вариаций, таким образом, существует концептуальный пробел относительно связи между мутацией и потенциальным изменением фенотипа. Чтобы мутация могла легко изменить фенотип и, следовательно, быть видимой для естественного отбора, она должна изменить онтогенетическую траекторию - процесс, называемый перепрограммированием развития. Некоторые виды перепрограммирования более вероятны, чем другие, учитывая природу карты генотип-фенотип, которая определяет склонность системы к изменению в определенном направлении, тем самым создавая предвзятость. Другими словами, лежащая в основе архитектура систем развития влияет на возможные фенотипические результаты.
Однако предвзятость в развитии может развиваться посредством естественного отбора, и оба процесса одновременно влияют на фенотипическую эволюцию. Например, смещение развития может повлиять на скорость или путь к адаптивному пику (фенотип высокой приспособленности), и, наоборот, сильный направленный отбор может изменить смещение развития, чтобы увеличить фенотипические вариации в направлении отбора. Таким образом, оба направления мысли дополняют друг друга и должны быть объединены, чтобы лучше понять процессы, лежащие в основе эволюционных изменений.
 Предвзятость в развитии для непрерывных персонажей. Если главная ось вариации (красные стрелки) ортогональна направлению выбора (пунктирная линия), ковариация признаков будет ограничивать адаптивную эволюцию. И наоборот, если главная ось вариации совпадает с направлением отбора, ковариация признаков будет способствовать адаптивной эволюции.
Предвзятость в развитии для непрерывных персонажей. Если главная ось вариации (красные стрелки) ортогональна направлению выбора (пунктирная линия), ковариация признаков будет ограничивать адаптивную эволюцию. И наоборот, если главная ось вариации совпадает с направлением отбора, ковариация признаков будет способствовать адаптивной эволюции.  Многомерное представление видов в морфопространстве. Каждая ось соответствует признаку, а точки соответствуют организмам с определенными комбинациями значений признаков. В этом случае оси представляют форму вида рыб.
Многомерное представление видов в морфопространстве. Каждая ось соответствует признаку, а точки соответствуют организмам с определенными комбинациями значений признаков. В этом случае оси представляют форму вида рыб. Морфопространство - это количественное представление фенотипов в многомерном пространстве, где каждое измерение соответствует признаку. Затем фенотип каждого организма или вида представлен в виде точки в этом пространстве, которая суммирует комбинацию значений или состояний по каждому конкретному признаку. Этот подход используется для изучения эволюции реализованных фенотипов по сравнению с теми, которые теоретически возможны, но не существуют.
Описание и понимание движущих сил распределения фенотипическая изменчивость в природе - одна из основных целей эволюционной биологии. Один из способов изучения распределения фенотипической изменчивости - это изображение объема морфопространства, занимаемого набором организмов или видов. Теоретически может существовать естественный процесс, который генерирует почти равномерный (квазистохастический) паттерн фенотипов в морфопространстве, с учетом того, что необходимые новые виды имеют тенденцию занимать точку в морфопространстве, близкую к таковым его филогенетических родственников. Однако в настоящее время широко признано, что организмы не распределены равномерно по морфопространству, т.е. изотропная изменчивость, а вместо этого распределена неслучайно, то есть анизотропная изменчивость. Другими словами, существует несоответствие между очевидными (или теоретическими) возможными фенотипами и их реальной доступностью.
 Онтогенетически невозможное существо
Онтогенетически невозможное существо Таким образом, некоторые фенотипы недоступны (или невозможны) из-за основной архитектуры траектории развития, а другие доступны (или возможны). Однако из возможных фенотипов некоторые «легче» или более вероятны, чем другие. Например, такой фенотип, как классическая фигура дракона (т. Е. Гигантского рептилийоподобного существа с двумя парами конечностей и передней парой крыльев), может быть невозможен, потому что у позвоночных передние конечности и передняя пара крыльев гомологичные символы (например, птицы и летучие мыши) и, таким образом, являются взаимоисключающими. С другой стороны, если два фенотипа возможны (и одинаково подходят), но для одной формы перепрограммирования требуется только одна мутация, а для другой - две или более, первое будет более вероятно (при условии, что генетические мутации происходят случайно).
Важное различие между структурализмом и функционализмом касается, прежде всего, интерпретации причин появления пустых областей в морфопространстве (то есть несуществующих фенотипов): с точки зрения функционализма, пустые пространства соответствуют фенотипам, которые являются одновременно онтогенетически возможны и равновероятны, но исключаются естественным отбором из-за их низкой приспособленности. Напротив, с точки зрения структуралистов, пустые пространства соответствуют онтогенетически невозможным или невероятным фенотипам, таким образом, подразумевая предвзятость в типах фенотипов, которые могут быть получены при условии равного количества вариаций (генетических мутаций) в обеих моделях.
 Изменения раковины в природе
Изменения раковины в природе На классическом естественном примере систематической ошибки было показано, что только небольшая часть всех возможных форм раковин улиток реализуется в природе, а реальные виды ограничиваются дискретными областями оболочка-морфопространство, а не непрерывное распространение. На другом природном примере было показано, что обитающие в почве многоножки имеют огромные различия в количестве пар ног, самая низкая - 27, а самая высокая - 191 пара; однако не существует видов с четным числом пар ног, что указывает на то, что либо эти фенотипы каким-то образом ограничиваются во время развития, либо существует стремление развития к нечетным числам. И наоборот, аномалии развития (или тератологии ) также использовались в качестве классического примера для понимания логики механизмов, вызывающих вариации. Например, у широкого круга животных, от рыб до людей, двуглавые организмы встречаются гораздо чаще, чем трехголовые; аналогично сиамские близнецы теоретически могут «слиться» через любую область тела, но слияние чаще происходит в области живота. Эта тенденция была названа трансвидовым параллелизмом, предполагая существование глубоких исторических правил, управляющих выражением аномальных форм у отдаленно родственных видов.
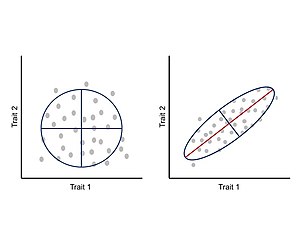 Представление отношения между двумя признаками. Слева: нет ковариации признаков. Каждая черта меняется независимо от другой. Справа: ковариация признака вызывает положительную корреляцию между признаками, где увеличение одного признака коррелирует с увеличением другого признака (ковариация также может давать отрицательную корреляцию). Красная линия внутри эллипса представляет собой главный собственный вектор матрицы вариации-ковариации.
Представление отношения между двумя признаками. Слева: нет ковариации признаков. Каждая черта меняется независимо от другой. Справа: ковариация признака вызывает положительную корреляцию между признаками, где увеличение одного признака коррелирует с увеличением другого признака (ковариация также может давать отрицательную корреляцию). Красная линия внутри эллипса представляет собой главный собственный вектор матрицы вариации-ковариации. Было высказано предположение, что интеграция или ковариация между признаками во время развития ограничивают фенотипическую эволюцию определенными областями морфопространства и ограничивают адаптивную эволюцию. Эти аллометрические изменения широко распространены по природе и могут объяснять большое разнообразие реализованных морфологий и последующих экологических и физиологических изменений. В рамках этого подхода фенотип рассматривается как интегрированная система, в которой каждый признак развивается и развивается вместе с другими признаками, и, таким образом, изменение одного признака коррелированно влияет на взаимодействующие части. Корреляция между признаками является следствием архитектуры карты генотип-фенотип, особенно плейотропных эффектов основных генов. Это коррелированное изменение между признаками можно измерить и проанализировать с помощью матрицы ковариации фенотипа (P-матрицы), которая суммирует размеры фенотипической изменчивости и главную ось вариации.
Количественная генетика - это статистическая основа, в основном занимающаяся моделированием эволюции непрерывных признаков. В рамках этой концепции корреляция между признаками может быть результатом двух процессов: 1) естественный отбор, действующий одновременно на несколько признаков, гарантирующий, что они наследуются вместе (т. Е. неравновесие по сцеплению ), или 2) естественный отбор, действующий на одно признак, вызывающий коррелированное изменение других признаков из-за плейотропных эффектов генов. Для набора признаков уравнение, описывающее дисперсию между признаками, представляет собой многомерное уравнение селекционера Δz = β x G, где Δz - вектор различий в средних признаках, β - вектор коэффициентов отбора, и G - матрица аддитивной генетической дисперсии и ковариации между признаками. Таким образом, непосредственная способность популяции реагировать на отбор определяется G-матрицей, в которой дисперсия является функцией постоянной генетической изменчивости, а ковариация возникает из-за плейотропии и неравновесия по сцеплению. Хотя G-матрица является одним из наиболее важных параметров для изучения эволюционируемости, мутационная матрица (M-матрица), также известная как распределение мутационных эффектов, имеет эквивалентное значение. M-матрица описывает потенциальные эффекты новых мутаций на существующие генетические вариации и ковариации, и эти эффекты будут зависеть от эпистатических и плейотропных взаимодействий лежащих в основе генов. Другими словами, M-матрица определяет G-матрицу и, таким образом, реакцию на выбор совокупности. Подобно P-матрице, G-матрица описывает главную ось вариации.
 Морфопространство и фитнес-ландшафт с одним оптимумом фитнеса. Для популяции, проходящей направленный отбор, главная ось вариации (наибольшая ось белого эллипса) будет смещать основное направление траектории в сторону оптимума приспособленности (стрелка). Скорость морфологического изменения будет обратно пропорциональна углу (бета), образованному между направлением выбора (пунктирная линия) и главной осью изменения.
Морфопространство и фитнес-ландшафт с одним оптимумом фитнеса. Для популяции, проходящей направленный отбор, главная ось вариации (наибольшая ось белого эллипса) будет смещать основное направление траектории в сторону оптимума приспособленности (стрелка). Скорость морфологического изменения будет обратно пропорциональна углу (бета), образованному между направлением выбора (пунктирная линия) и главной осью изменения. Общее следствие P-матриц и G-матриц заключается в том, что эволюция будут стремиться следовать «путем наименьшего сопротивления». Другими словами, если основная ось вариации совпадает с направлением отбора, ковариация (генетическая или фенотипическая) будет способствовать скорости адаптивной эволюции; однако, если главная ось изменения ортогональна направлению выбора, ковариация будет ограничивать скорость адаптивной эволюции. В целом, для популяции, находящейся под влиянием единственного оптимума приспособленности, скорость морфологической дивергенции (от предкового к новому фенотипу или между парами видов ) обратно пропорциональна углу, образуемому основным ось вариации и направление выбора, вызывая искривленную траекторию через морфопространство.
Из P-матрицы для набора символов могут быть извлечены два важных показателя склонности к изменению: 1) Ответственность : способность системы развития изменяться в любом направлении; и 2) эволюционируемость : способность системы развития изменяться в направлении естественного отбора. В последнем случае основная ось фенотипической изменчивости совпадает с направлением отбора. Точно так же из G-матрицы наиболее важным параметром, описывающим склонность к изменению, является опережающий собственный вектор G (g max), который описывает направление наибольшего аддитивная генетическая дисперсия для набора непрерывных признаков в популяциях. Для популяции, проходящей направленный отбор, g max будет смещать основное направление траектории.
 Различные виды позвоночных развили меланические формы в результате параллельных мутаций в гене mc1r.
Различные виды позвоночных развили меланические формы в результате параллельных мутаций в гене mc1r.GRN представляют собой модульные, многослойные и полуиерархические системы генов и их продуктов: каждая фактор транскрипции обеспечивает множественные входные данные для других генов, создавая сложный набор взаимодействий, и информация, касающаяся времени, места и количества экспрессии генов, обычно поступает от нескольких высокоуровневых контрольных генов через несколько промежуточных генов к периферийным генным батареям. которые в конечном итоге определяют судьбу каждой клетки. Этот тип архитектуры подразумевает, что гены контроля высокого уровня имеют тенденцию быть более плейотропными, влияя на несколько нижестоящих генов, тогда как промежуточные и периферические гены имеют тенденцию иметь умеренный или низкий плейотропный эффект, соответственно.
В В целом ожидается, что вновь возникшие мутации с более высоким доминированием и меньшим количеством плейотропных и эпистатических эффектов с большей вероятностью станут мишенями эволюции, таким образом, иерархическая архитектура путей развития может искажать генетическую основу эволюционных изменений. Например, ожидается, что гены внутри GRN с «оптимально плейотропными» эффектами, то есть гены, которые оказывают наиболее широкое влияние на отборный признак, но мало влияют на другие признаки, будут накапливать большую долю мутаций, вызывающих эволюционные изменения. Эти стратегически расположенные гены обладают потенциалом фильтровать случайные генетические вариации и переводить их в неслучайно функционально интегрированные фенотипы, делая адаптивные варианты эффективно доступными для отбора, и, таким образом, многие мутации, способствующие фенотипической эволюции, могут быть сконцентрированы в этих генах. 49>
Перспектива карты генотип-фенотип устанавливает, что способ, которым генотипические вариации могут быть сопоставлены с фенотипическими вариациями, имеет решающее значение для способности системы развиваться. Преобладание нейтральных мутаций в природе означает, что биологические системы имеют больше генотипов, чем фенотипов, и как следствие этой связи «многие к немногим» между генотипами а фенотип - это наличие нейтральных сетей. В процессе развития нейтральные сети представляют собой кластеры GRN, которые различаются только одним взаимодействием между двумя узлами (например, заменой транскрипции супрессией) и, тем не менее, дают одинаковый фенотипический результат. В этом смысле индивидуальный фенотип в популяции может быть сопоставлен с несколькими эквивалентными GRN, которые вместе составляют нейтральную сеть. И наоборот, GRN, который отличается одним взаимодействием и вызывает другой фенотип, считается ненейтральным. Учитывая эту архитектуру, вероятность мутации от одного фенотипа к другому будет зависеть от количества нейтральных соседей по сравнению с ненейтральными соседями для конкретной GRN, и, таким образом, на фенотипические изменения будет влиять положение GRN в сети. и будет склоняться к изменениям, требующим нескольких мутаций для достижения соседнего ненейтрального GRN.