
Войти
 Структура генной регуляторной сети
Структура генной регуляторной сети  Процесс управления генной регуляторной сетью
Процесс управления генной регуляторной сетью A ген (или генетическая ) регуляторная сеть (GRN ) - это совокупность молекулярных регуляторов, которые взаимодействуют друг с другом и с другими веществами в клетке, чтобы управлять экспрессией гена уровни мРНК и белков. Они играют центральную роль в морфогенезе, создании структур тела, что, в свою очередь, является центральным в эволюционной биологии развития (evo-DevO).
Регулятор может представлять собой ДНК, РНК, белок и их комплексы. Взаимодействие может быть прямым или непрямым (через транскрибируемую РНК или транслируемый белок). В общем, каждая молекула мРНК производит определенный белок (или набор белков). В некоторых случаях этот белок будет структурным и будет накапливаться на клеточной мембране или внутри клетки, придавая ей особые структурные свойства. В других случаях белок будет ферментом, то есть микромашиной, которая катализирует определенную реакцию, например, расщепление источника пищи или токсина. Некоторые белки служат только для активации других генов, и это факторы транскрипции, которые являются основными участниками регуляторных сетей или каскадов. Связываясь с областью промотора в начале других генов, они включают их, инициируя продукцию другого белка и т. Д. Некоторые факторы транскрипции являются ингибирующими.
В одноклеточных организмах регуляторные сети реагируют на внешнюю среду, оптимизируя клетку в данный момент времени для выживания в этой среде. Таким образом, дрожжевая клетка, оказавшись в растворе сахара, включает гены, вырабатывающие ферменты, которые превращают сахар в спирт. Этот процесс, который мы связываем с виноделием, заключается в том, как дрожжевые клетки зарабатывают себе на жизнь, получая энергию для размножения, что при нормальных обстоятельствах может улучшить ее шансы на выживание.
У многоклеточных животных тот же принцип был поставлен на службу генным каскадам, контролирующим форму тела. Каждый раз, когда клетка делится, возникают две клетки, которые, хотя и содержат один и тот же геном полностью, могут различаться по тому, какие гены включаются и производят белки. Иногда «самоподдерживающаяся петля обратной связи» гарантирует, что клетка сохраняет свою идентичность и передает ее. Менее понятен механизм эпигенетики, с помощью которого модификация хроматина может обеспечивать клеточную память путем блокирования или разрешения транскрипции. Основной особенностью многоклеточных животных является использование градиентов морфогена, которые, по сути, обеспечивают систему позиционирования, которая сообщает клетке, где она находится в теле и, следовательно, какой клеткой ей стать. Ген, который включен в одной клетке, может производить продукт, который покидает клетку и распространяется через соседние клетки, проникая в них и включая гены, только когда он присутствует выше определенного порогового уровня. Таким образом, в этих клетках индуцируется новая судьба, и они могут даже генерировать другие морфогены, которые возвращают сигнал исходной клетке. На больших расстояниях морфогены могут использовать активный процесс передачи сигнала. Такая передача сигналов контролирует эмбриогенез, построение плана тела с нуля посредством серии последовательных шагов. Они также контролируют и поддерживают взрослые тела посредством процессов обратной связи, и потеря такой обратной связи из-за мутации может быть ответственной за пролиферацию клеток, которая наблюдается при раке. Параллельно с этим процессом построения структуры каскад генов включает гены, которые создают структурные белки, которые придают каждой клетке необходимые ей физические свойства.
С одной стороны, биологические клетки можно рассматривать как «частично смешанные пакеты» биологических химикатов - при обсуждении сетей регуляции генов эти химические вещества в основном являются информационные РНК (мРНК) и белки, которые возникают в результате экспрессии генов. Эти мРНК и белки взаимодействуют друг с другом с разной степенью специфичности. Некоторые распространяются по камере. Другие связаны с клеточными мембранами, взаимодействуя с молекулами окружающей среды. Третьи проходят через клеточные мембраны и передают сигналы дальнего действия другим клеткам многоклеточного организма. Эти молекулы и их взаимодействия составляют регуляторную сеть генов. Типичная регуляторная сеть генов выглядит примерно так:
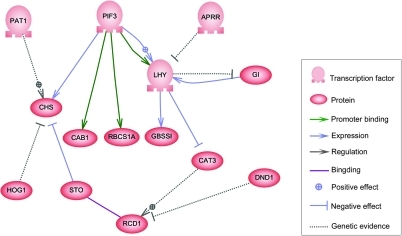 Пример регуляторной сети
Пример регуляторной сети Узлы этой сети могут представлять гены, белки, мРНК, комплексы белок / белок или клеточные процессы. Узлы, которые изображены лежащими вдоль вертикальных линий, связаны с интерфейсами ячейки / среды, в то время как остальные являются свободно плавающими и могут распространяться. Ребра между узлами представляют собой взаимодействия между узлами, которые могут соответствовать индивидуальным молекулярным реакциям между ДНК, мРНК, миРНК, белками или молекулярными процессами, посредством которых продукты одного гена влияют на продукты другого, хотя отсутствие экспериментально полученной информации часто означает, что некоторые реакции не моделируются на таком высоком уровне детализации. Эти взаимодействия могут быть индуктивными (обычно представлены стрелками или знаком +), с увеличением концентрации одного, ведущим к увеличению другого, ингибирующим (представлены закрашенными кружками, тупыми стрелками или знаком минус), с увеличением в одном ведет к уменьшению в другом, или в двойном, когда в зависимости от обстоятельств регулятор может активировать или ингибировать целевой узел. Узлы могут регулировать себя прямо или косвенно, создавая петли обратной связи, которые образуют циклические цепочки зависимостей в топологической сети. Сетевая структура - это абстракция молекулярной или химической динамики системы, описывающая множество способов, которыми одно вещество влияет на все другие, с которыми оно связано. На практике такие GRN выводятся из биологической литературы по данной системе и представляют собой квинтэссенцию коллективных знаний о наборе связанных биохимических реакций. Чтобы ускорить ручное курирование GRN, некоторые недавние попытки пытаются использовать для этой цели анализ текста, курируемые базы данных, сетевой вывод из массивных данных, проверку моделей и другие технологии извлечения информации.
Гены можно рассматривать как узлы в сети, где входными данными являются белки, такие как факторы транскрипции, а выходными данными - уровень экспрессии генов. Значение узла зависит от функции, которая зависит от значения его регуляторов на предыдущих временных шагах (в описанной ниже логической сети это логические функции, обычно И, ИЛИ и НЕ). Эти функции интерпретировались как выполнение своего рода обработки информации внутри ячейки, которая определяет поведение ячейки. Основными драйверами внутри клетки являются концентрации некоторых белков, которые определяют как пространственные (расположение в клетке или ткани), так и временные (клеточный цикл или стадия развития) координаты клетки, как своего рода «клеточную память». Генные сети только начинают понимать, и это следующий шаг для биологии, чтобы попытаться вывести функции для каждого генного «узла», чтобы помочь понять поведение системы на возрастающих уровнях сложности., от гена к сигнальному пути, клеточному или тканевому уровню.
Математические модели GRN были разработаны, чтобы фиксировать поведение моделируемой системы и в некоторых случаях генерировать прогнозы, соответствующие экспериментальным наблюдениям. В некоторых других случаях доказано, что модели делают точные новые прогнозы, которые можно проверить экспериментально, предлагая, таким образом, новые подходы для исследования в эксперименте, которые иногда не учитывались бы при разработке протокола экспериментальной лаборатории. Методы моделирования включают дифференциальные уравнения (ODE), булевы сети, сети Петри, байесовские сети, графические модели гауссовских сетей, Стохастический и Обработка вычислений. И наоборот, были предложены методы для создания моделей GRN, которые лучше всего объясняют набор временных рядов наблюдений. Недавно было показано, что сигнал ChIP-seq модификации гистона больше коррелирует с мотивами факторов транскрипции на промоторах по сравнению с уровнем РНК. Следовательно, предполагается, что модификация гистонов временного ряда ChIP-seq может обеспечить более надежный вывод о ген-регуляторных сетях по сравнению с методами, основанными на уровнях экспрессии.
Обычно считается, что сети регуляторов генов состоят из нескольких тесно связанных узлов (концентраторов ) и множество плохо связанных узлов, вложенных в иерархический режим регулирования. Таким образом, сети регуляции генов приближаются к топологии иерархической сети без масштабирования. Это согласуется с мнением, что большинство генов обладают ограниченной плейотропией и действуют в рамках регуляторных модулей. Считается, что эта структура развивается благодаря предпочтительному прикреплению дублированных генов к более тесно связанным генам. Недавние исследования также показали, что естественный отбор имеет тенденцию отдавать предпочтение сетям с разреженной связностью.
В основном сети могут развиваться двумя способами, причем оба могут происходить одновременно. Во-первых, топология сети может быть изменена путем добавления или вычитания узлов (генов) или частей сети (модулей), которые могут быть выражены в разных контекстах. Drosophila сигнальный путь Hippo является хорошим примером. Сигнальный путь Hippo контролирует как митотический рост, так и постмитотическую клеточную дифференцировку. Недавно было обнаружено, что сеть, в которой работает сигнальный путь Hippo, различается между этими двумя функциями, что, в свою очередь, изменяет поведение сигнального пути Hippo. Это предполагает, что сигнальный путь Hippo работает как консервативный регуляторный модуль, который может использоваться для множества функций в зависимости от контекста. Таким образом, изменение топологии сети может позволить сохраняемому модулю выполнять несколько функций и изменять конечный результат работы сети. Второй способ развития сетей - это изменение силы взаимодействий между узлами, например, насколько сильно фактор транскрипции может связываться с цис-регуляторным элементом. Было показано, что такие различия в силе краев сети лежат в основе межвидовых вариаций в формировании паттерна судьбы клеток вульвы Caenorhabditis червей.
 Петля прямой связи
Петля прямой связи Другой широко распространенный Цитируемой характеристикой регуляторной сети генов является обилие определенных повторяющихся подсетей, известных как сетевые мотивы. Сетевые мотивы можно рассматривать как повторяющиеся топологические паттерны при разделении большой сети на маленькие блоки. Предыдущий анализ выявил несколько типов мотивов, которые чаще появляются в сетях регуляции генов, чем в случайно сгенерированных сетях. Например, один из таких мотивов называется петлями прямой связи, которые состоят из трех узлов. Этот мотив является наиболее распространенным среди всех возможных мотивов, состоящих из трех узлов, как показано в регуляторных сетях генов мух, нематод и человека.
Было предложено, чтобы обогащенные мотивы следовали за конвергентными evolution, предполагая, что это «оптимальные конструкции» для определенных нормативных целей. Например, моделирование показывает, что петли с прямой связью способны координировать изменение в узле A (с точки зрения концентрации и активности) и динамику выражения узла C, создавая различные режимы ввода-вывода. Система использования галактозы в E. coli содержит петлю с прямой связью, которая ускоряет активацию утилизации галактозы оперон galETK, потенциально способствуя метаболическому переходу в галактозу при истощении глюкозы. Петля с прямой связью в системах утилизации арабинозы E.coli задерживает активацию оперона катаболизма арабинозы и транспортеров, потенциально избегая ненужных метаболических переходов из-за временных флуктуаций восходящих сигнальных путей. Аналогично в сигнальном пути Wnt Xenopus петля прямой связи действует как детектор кратного изменения, который реагирует на кратное изменение, а не на абсолютное изменение уровня β-катенина, потенциально увеличивая устойчивость к колебаниям уровня β-катенина. Следуя гипотезе конвергентной эволюции, обогащение контуров прямой связи будет адаптацией для быстрого отклика и устойчивости к шуму. Недавнее исследование показало, что дрожжи, выращенные в среде с постоянным содержанием глюкозы, развивают мутации в путях передачи сигналов глюкозы и в путях регуляции роста, что позволяет предположить, что регуляторные компоненты, реагирующие на изменения окружающей среды, не требуются при постоянной среде.
С другой стороны, некоторые исследователи выдвинуть гипотезу о том, что обогащение сетевых мотивов не адаптивно. Другими словами, сети регуляции генов могут развиваться до аналогичной структуры без специального выбора предлагаемого поведения ввода-вывода. Поддержка этой гипотезы часто исходит из компьютерного моделирования. Например, колебания в количестве петель прямой связи в модели, которая имитирует эволюцию регуляторных сетей генов путем случайного перенастройки узлов, может указывать на то, что обогащение петель прямой связи является побочным эффектом эволюции. В другой модели эволюции сетей регуляторов генов соотношение частот дупликации и удаления генов оказывает большое влияние на топологию сети: определенные соотношения приводят к обогащению петель прямой связи и создают сети, которые демонстрируют особенности иерархических сетей без масштабов. De novo эволюция когерентных контуров прямой связи типа 1 была продемонстрирована с помощью вычислений в ответ на выбор их гипотетической функции фильтрации короткого паразитного сигнала, поддерживающей адаптивную эволюцию, но для неидеализированного шума - динамическую систему подачи Вместо этого предпочтение было отдано прямому регулированию с другой топологией.
Регуляторные сети позволяют бактериям адаптироваться практически к любой экологической нише на Земле. Сеть взаимодействий между различными типами молекул, включая ДНК, РНК, белки и метаболиты, используется бактериями для достижения регуляции экспрессии генов. У бактерий основная функция регулирующих сетей состоит в том, чтобы контролировать реакцию на изменения окружающей среды, например, на состояние питания и экологический стресс. Сложная организация сетей позволяет микроорганизмам координировать и интегрировать несколько сигналов окружающей среды.
Обычно такую сеть моделируют с помощью набора связанных обыкновенных дифференциальных уравнений (ODE) или SDE, описывающих кинетику реакции составляющих частей. Предположим, что наша нормативная сеть имеет 



, где функции 

Решая для фиксированной точки системы:

для всех 
В следующем примере показано, как логическая сеть может моделировать GRN вместе с его генными продуктами (выходами) и веществами из окружающей среды, которые влияют на него (входы). Стюарт Кауфман был одним из первых биологов, использовавших метафору булевых сетей для моделирования генетических регуляторных сетей.
Достоверность модели можно проверить, сравнив результаты моделирования со временем серия наблюдений. Частичная проверка булевой сетевой модели также может происходить из проверки предсказанного существования еще неизвестной регуляторной связи между двумя конкретными факторами транскрипции, каждый из которых является узлами модели.
Непрерывные сетевые модели GRN являются расширением булевых сетей, описанных выше. Узлы по-прежнему представляют собой гены и связи между ними, регулирующие влияние на экспрессию генов. Гены в биологических системах демонстрируют непрерывный диапазон уровней активности, и утверждалось, что использование непрерывного представления захватывает несколько свойств регуляторных сетей генов, не представленных в булевой модели. Формально большинство этих подходов похожи на искусственную нейронную сеть, поскольку входные данные узла суммируются, а результат служит входом для сигмовидной функции, например, но белки часто контролируют экспрессию генов в синергетическом режиме., т.е. нелинейным, способом. Однако теперь существует непрерывная сетевая модель, которая позволяет группировать входные данные для узла, тем самым реализуя другой уровень регулирования. Эта модель формально ближе к рекуррентной нейронной сети более высокого порядка . Эта же модель также использовалась для имитации эволюции клеточной дифференциации и даже многоклеточного морфогенеза.
Недавние экспериментальные результаты продемонстрировали, что экспрессия генов является стохастической. процесс. Таким образом, многие авторы сейчас используют стохастический формализм после работы Аркина и др. Работы по экспрессии одного гена и небольших синтетических генетических сетях, таких как генетический тумблер Тима Гарднера и Джим Коллинз, предоставили дополнительные экспериментальные данные о фенотипической изменчивости и стохастической природе экспрессии генов. Первые версии стохастических моделей экспрессии генов включали только мгновенные реакции и управлялись алгоритмом Гиллеспи.
, поскольку некоторые процессы, такие как транскрипция генов, включают множество реакций и не могли быть правильно смоделированы как мгновенная реакция в одноэтапный, было предложено моделировать эти реакции как одноэтапные множественные отложенные реакции, чтобы учесть время, необходимое для завершения всего процесса.
Отсюда был предложен набор реакций, которые позволяют генерирование GRN. Затем они моделируются с использованием модифицированной версии алгоритма Гиллеспи, который может моделировать несколько реакций с задержкой по времени (химические реакции, в которых каждому из продуктов предоставляется временная задержка, определяющая, когда он будет выпущен в систему в качестве «готового продукта»)..
Например, основная транскрипция гена может быть представлена следующей одностадийной реакцией (RNAP - это РНК-полимераза, RBS - сайт связывания РНК-рибосомы, а Pro i - это промоторная область гена i):

Кроме того, кажется, существует компромисс между шумом в экспрессии генов, скоростью, с которой гены могут переключаться, и метаболическими издержками, связанными с их функционированием. Более конкретно, для любого заданного уровня метаболических затрат существует оптимальный компромисс между шумом и скоростью обработки, а увеличение метаболических затрат приводит к лучшему компромиссу между скоростью и шумом.
В недавней работе был предложен симулятор (SGNSim, имитатор стохастических генных сетей), который может моделировать GRN, где транскрипция и трансляция моделируются как события с множественной задержкой по времени, а ее динамика управляется алгоритмом стохастического моделирования (SSA), способным работать с событиями с множественной задержкой по времени. Временные задержки могут быть получены из нескольких распределений, а скорости реакции - из сложных функций или физических параметров. SGNSim может генерировать ансамбли GRN в пределах набора определяемых пользователем параметров, таких как топология. Его также можно использовать для моделирования конкретных GRN и систем химических реакций. Также можно моделировать генетические нарушения, такие как делеции генов, сверхэкспрессия генов, вставки, мутации сдвига рамки считывания.
GRN создается из графа с желаемой топологией с наложением внутренних и внешних распределений. На активность промоторов генов влияют продукты экспрессии других генов, которые действуют как входы, в форме мономеров или объединены в мультимеры и считаются прямыми или косвенными. Затем каждый прямой ввод назначается сайту оператора, и различным факторам транскрипции может быть разрешено или нет конкурировать за один и тот же сайт оператора, в то время как косвенные входы задаются целью. Наконец, каждому гену назначается функция, определяющая ответ гена на комбинацию факторов транскрипции (состояние промотора). Передаточные функции (то есть то, как гены реагируют на комбинацию входных сигналов) могут быть назначены каждой комбинации состояний промотора по желанию.
В другой недавней работе были разработаны многомасштабные модели сетей регуляции генов, которые сосредоточены на приложениях синтетической биологии. Были использованы симуляции, моделирующие все биомолекулярные взаимодействия в транскрипции, трансляции, регуляции и индукции регуляторных сетей генов, определяющие дизайн синтетических систем.
Другая работа была сосредоточена на предсказании уровни экспрессии генов в регуляторной сети генов. Подходы, используемые для моделирования сетей регуляции генов, были ограничены, чтобы их можно было интерпретировать, и, как следствие, они обычно являются упрощенными версиями сети. Например, булевы сети использовались из-за их простоты и способности обрабатывать зашумленные данные, но терять информацию о данных из-за двоичного представления генов. Кроме того, искусственные нейронные сети не используют скрытый слой, чтобы их можно было интерпретировать, теряя возможность моделировать корреляции более высокого порядка в данных. Используя модель, которую нельзя интерпретировать, можно создать более точную модель. Возможность более точно предсказать экспрессию генов дает возможность исследовать, как лекарства влияют на систему генов, а также определять, какие гены взаимосвязаны в процессе. Этому способствовал конкурс DREAM, который способствует конкуренции за лучшие алгоритмы прогнозирования. В некоторых других недавних работах использовались искусственные нейронные сети со скрытым слоем.
Существует три класса рассеянного склероза: рецидивирующий-ремиттирующий (RRMS), первично-прогрессивный (PPMS) и вторично-прогрессивный (SPMS). Генная регуляторная сеть (GRN) играет жизненно важную роль в понимании механизма заболевания в этих трех различных классах рассеянного склероза.