
Войти
| Chr 22: 38,52 - 38,57 Мб | Chr 15: 79,56 - 79,61 Мб | ||||||
| PubMed поиск | |||||||
| Викиданные | |||||||
| |||||||
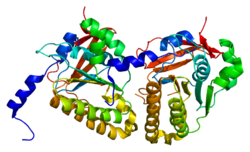
Гомолог белка мейотической рекомбинации DMC1 / LIM15 представляет собой белок, который у человека кодируется геном DMC1 .
Мейотический рекомбинационный белок Dmc1 является гомологом бактериального белка обмена цепей RecA. Dmc1 играет центральную роль в гомологичной рекомбинации в мейозе, собираясь на сайтах запрограммированных двухцепочечных разрывов ДНК и выполняя поиск аллельных последовательностей ДНК, расположенных на гомологичных хроматидах. Название «Dmc» означает «нарушенная мейотическая кДНК» и относится к методу, использованному для ее открытия, который включал использование клонов из библиотеки кДНК, специфичной для мейоза, для прямого нокаута мутаций в большом количестве экспрессируемых мейотических генов. Белок Dmc1 является одним из двух гомологов RecA, обнаруженных в эукариотических клетках, другим является Rad51. У почкующихся дрожжей Rad51 служит белком обмена цепей в митозе, где он важен для восстановления разрывов ДНК. Rad51 превращается в дополнительный фактор для Dmc1 во время мейоза за счет ингибирования его активности обмена цепей. Гомологи DMC1 были идентифицированы у многих организмов, включая дивергентные грибы, растения и млекопитающих, включая человека.
Ген и белок DMC1 были обнаружены в почкующихся дрожжах S. cerevisiae, когда он работал научным сотрудником в лаборатории Нэнси Клекнер в Гарвардском университете.
Белок, кодируемый этим геном, необходим для мейотической гомологичной рекомбинации. Генетическая рекомбинация в мейозе играет важную роль в создании разнообразия генетической информации и способствует редукционной сегрегации хромосом, которая должна происходить для образования гамет во время полового размножения.
Подобно другим членам семейства Rad51 / RecA, Dmc1 стабилизирует промежуточные звенья обмена цепей (Rad1 / RecA-растянутая ДНК или RS-ДНК) в растянутых триплетах, подобных B-форме ДНК. Каждая молекула белка связывает триплет нуклеотидов, и сила этого связывания, оцениваемая по изменению свободной энергии Гиббса, может быть оценена по продолжительности времени, в течение которого меченый зонд дцДНК с коротким гомологичная последовательность остается связанной с ДНК, содержащей короткий участок гомологии с ней. Исследование этого типа показало, что несоответствие в любом из трех положений в конце участка гомологии не увеличит продолжительность времени, в течение которого зонд остается связанным, а в конструкциях Rad51 или RecA внутреннее несоответствие вызовет аналогичное сокращение времени связывания. Все ферменты способны «перешагнуть» несоответствие и продолжить связывать зонд более прочно, если существует более протяженная область гомологии. Однако с Dmc1 триплет с одним внутренним (но не концевым) несоответствием будет вносить вклад в стабильность связывания зонда в такой же степени, как и триплет без несоответствия. Таким образом, Dmc1 особенно подходит для выполнения своей роли специфичной для мейоза рекомбиназы, поскольку эта активность позволяет ему более эффективно катализировать рекомбинацию между последовательностями, которые не полностью совпадают.
DMC1 Было показано, что (ген) взаимодействует с RAD51. Было также показано, что белок связывает Tid1 (Rdh54), Mei5 / Sae3 и Hop2 / Mnd1. Все эти взаимодействующие белки действуют, усиливая активность Dmc1 в очищенных системах, а также считаются необходимыми для функции Dmc1 в клетках.
Во время мейоза две рекомбиназы, Rad51 и Dmc1 взаимодействуют с одноцепочечными ДНК для образования специализированных нитей, которые адаптированы для облегчения рекомбинации между гомологичными хромосомами. И Dmc1, и Rad51 обладают внутренней способностью к самоагрегированию. Присутствие нитей Rad51 стабилизирует соседние филаменты Dmc1 и, наоборот, Dmc1 стабилизирует соседние филаменты Rad51. Была предложена модель, в которой Dmc1 и Rad51 образуют отдельные филаменты на одной и той же одноцепочечной ДНК, и перекрестные помехи между двумя рекомбиназами влияют на их биохимические свойства.
Во время мейоза, даже в отсутствие активности обмена цепей Rad51, Dmc1, по-видимому, способен восстанавливать все разрывы мейотической ДНК, и это отсутствие не влияет на скорость кроссинговера мейоза.




 ..
.. 